本章重點摘要:
主題 1. 演替的基本概念
A. 18世紀末期由歐洲開始發展
B. 英文名詞為succession,高中以下常翻譯為”消長”
C. 定義
D. 先驅與極峰(極盛相)物種
E. 初級與次級演替
F. 基本模型的問題:時間尺度、干擾、單方向性
主題 2. 森林動態 (forest dynamics)、更新(recruitment)
A. 演替的動力來自於干擾
B. 干擾可發生在任何階段
C. 干擾發生時間與規模是主要的影響因子
D. 干擾若持續存在,不易演替至極盛相植群 –> 潛在植被
E. 理想上可以達到一個循環式的平衡 -> 動態平衡 (但實際很難)
F. 森林動態研究的發展與探討全球變遷議題有密切關聯
主題 3. 樹冠孔隙 (canopy gap)
A. 先驅相–極盛相樹種 -> 陽性樹種–陰性樹種 -> 不耐陰樹種—耐陰樹種
B. 徑級結構與樹種的耐陰性
C. 種子大小通常和休眠能力有關,能夠休眠的種子才能夠進入土壤種子庫
D. 孔隙是森林動態變化的驅動因子
E. 孔隙大小造成不同的干擾規模,進而影響森林動態
F. 孔隙提供不耐陰樹種存活的機會 -> 物種豐富度的增加
G. 孔隙扮演維持森林多樣性的重要角色
主題 4. 地被與土壤種子庫
A. 地被是森林更新一個極為關鍵的位置 -> 大型草本植物影響大樹小苗,大樹影響大型草本植物
B. 土壤種子庫往往不是直接補充耐陰樹種,而是加快不耐陰樹種在孔隙建立的速度 (注意多數植物一年只有一次果熟期)
主題 5. 森林動態觀念在森林復育的疑問
A. 是否需要根據演替序列進行植栽?
B. 如何達成樹種多樣性?
這章的主題是「森林動態」(Forest Dynamics)。介紹森林動態主要是讓大家知道:森林是活著的生物組成的,包括樹在內、植物在內,都是活著的,所以它會一直在改變。但是這改變是什麼,我們其實不用太害怕,比如說看到這森林裡面,有些大樹死了或怎樣,我們可能會擔心。其實有一些狀況,你要長期去看,才會知道它實際上是怎麼回事。森林動態這一章是要介紹這種觀念。
森林動態這章有五個主題,第一個主題介紹「演替」的觀念,在講到森林動態之前,都要回歸到「演替」。
主題 1. 演替的基本觀念
A.18世紀末期由歐洲開始發展
「演替」的觀念在現在生態學是非常基本的常識。但是「演替」是在18世紀從歐洲才開始發展,就基本知識來講,「演替」是一個滿新的觀念。
當時為什麼歐洲他們會注意到這件事情?因為早期歐洲他們有很廣大的雲杉林,人們一開始就好奇,想要了解這些雲杉林會怎麼樣,於是後來知道雲杉可以拿來做木材、或者了解有什麼物種跟雲杉在一起。久了當這些事情變成基本知識的時候,人類的思想的野心就會希望知道:這個森林它是怎麼來的,它會演變成什麼。這樣的思考點就讓人們去定期觀察這個森林的變化,慢慢地就出現所謂「演替」的觀念。
B.英文名詞為 succession,高中以下常翻譯為”消長”
演替的英文是「succession」,在大學課程裡面「succession」的中文翻譯為「演替」,可是高中以下的教科書大部分是譯為「消長」,所以大一學生一進大學聽到「演替」這個字眼,會愣在那邊不知道我們在講什麼。我就換個角度說,你們以前的課本可能講消長,他們就懂了。對一般的人來講,比較知道的是消長這個字,但是在英文是同一個字「succession」。
為什麼在大學傾向於用演替,而不是用消長?原因是-消長不是一個很好的名詞。消長的意思是:你消我長,某些東西可能弱勢了,然後有些東西就出現了,會變成一個非A即B的觀念。但是「演替」的觀念其實在講-怎麼樣去循環,「演替」已經講到「森林動態」、「森林更新」這一個觀念。
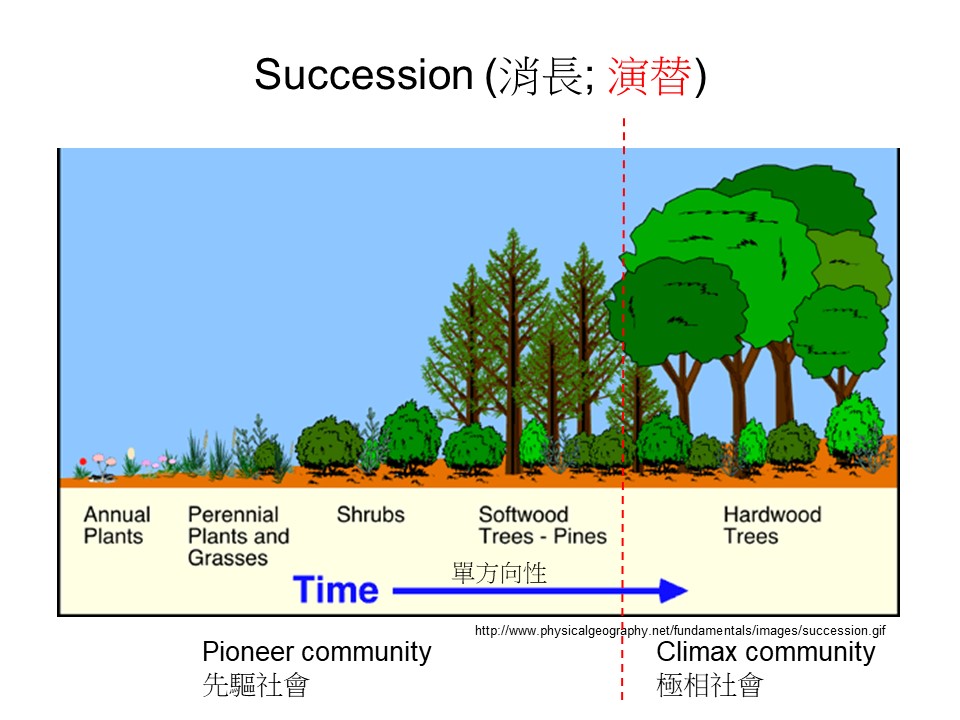
這是我們在生態學教科書最常用的演替圖。如果是一般高中以下的教科書大概都會用類似的圖,還有一些圖可能是畫一個湖泊,湖泊陸化之後慢慢有草、慢慢有樹這樣。
這張演替的基本概念圖想要表達的是:隨著時間的進程,一開始這個地方可能會長一些草本植物,這些草本植物可能是一年生的(Annual Plants),慢慢地有多年生的草本進來(Perennial Plants and Grasses),然後有一些小灌木進來,之後就變成一些灌叢(Shrubs),再變成一些森林。這個圖把森林分成兩類,一類是裸子植物的這種針葉樹林(Softwood Trees – Pines),然後慢慢演變成另一類被子植物的闊葉樹林(Hardwood Trees)。
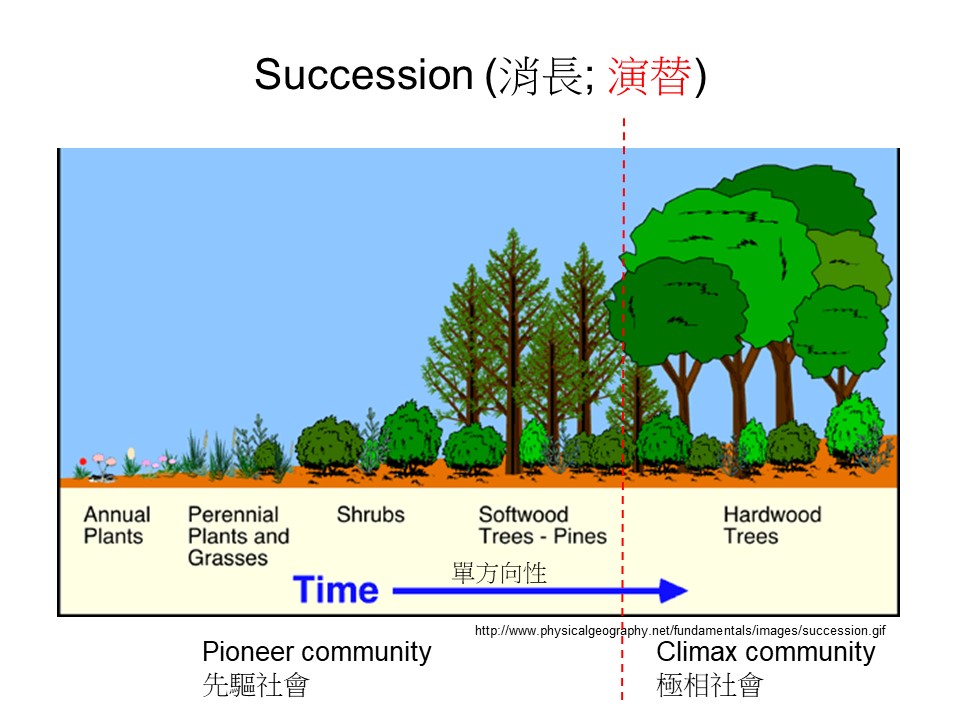
來源:Pidwirny, M. (2006). “Plant Succession”. Fundamentals of Physical Geography, 2nd Edition. 2022/12/15,http://www.physicalgeography.net/fundamentals/9i.html
這個示意圖表達的只是一種概念,不是真的發生在地球上的事情。生態學這種知識很複雜,因此我們就要把它簡單化,不然沒辦法濃縮到教科書裡。所以這張演替圖很簡單地講,如果今天有一塊地空出來,它會怎麼樣去變化,可是這張圖把演替的過程單純化,這是所謂的「想像的概念」,真實的演替不會這樣子運作。大家要把這事情釐清,我們在野外幾乎不會直接就看到這樣子的演變。
我希望盡可能把演替的觀念單純化,因為這比介紹植群、物種這些觀念要更學術一些,大家還是要盡可能理解,因為森林動態跟我們的森林復育很有關係。如果我們能夠抓到我們區域的、地方性的森林的演替脈絡的話,在做復育的時候才能夠做正確的選擇。
C.定義
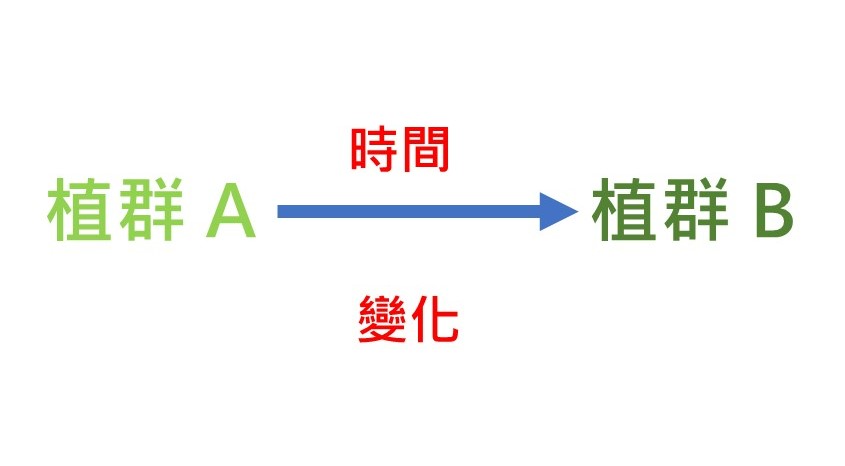
我們來思考一下演替的定義。這個演替基本模型圖牽涉到兩個核心概念:第一個核心概念是「植群」,因為演替是在講植群的演替。植群的定義是:一個地方在同一個時間的所有活著的植物的組合。我們在第一章植群的基礎觀念裡開宗明義講「植群是什麼?」,所下的定義就是:在特定的區域裡面,特定時間,所有活著的植物的組合,這個叫做「植群」。所以植群是演替的基本核心。
演替基本模型圖的第二個核心概念是「時間」。因為植群會變,為什麼會變?理由很簡單,植物是活的生物,它會有生老病死,所以在這個時間點它是這樣的植群,在某個時間點它可能不是這樣的植群,所以演替的另外一個動力就是在於「時間」。
如右圖所示,我們把一開始的植群定義為植群A,經過某些時間,經過某些變化,成為植群B,這就是演替的定義。
我們必須要強調,這個植群它代表的是「特定範圍內」的植群,不能把範圍改變,一定要探討同一個範圍這樣才有意義。所以在這個特定範圍裡面的植群,它隨著時間慢慢改變之後,它變成另外一個植群,這個我們稱為「演替」。這就是我們對演替所下的一個定義。
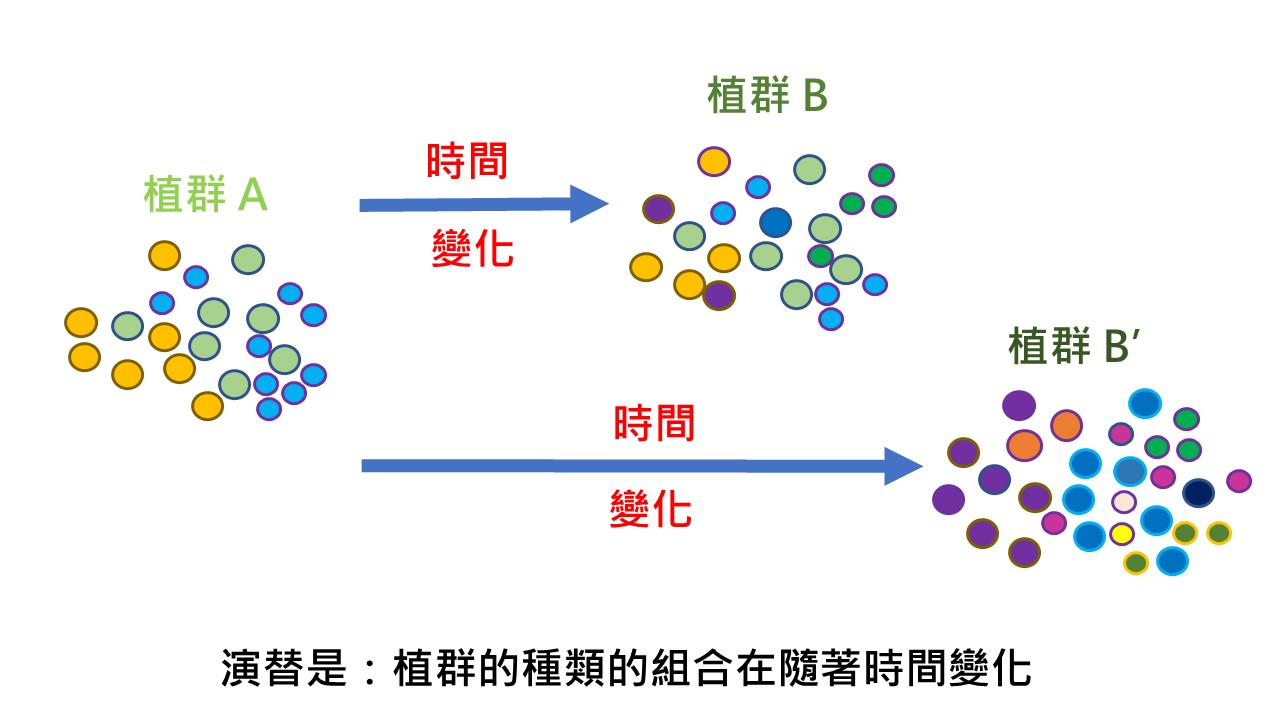
因為植群是植物物種的組合,當植群A變化到植群B的時候,當時間不是很久的話,植群A必然會有一些殘留。如果時間很久遠,可能植群A就全部換掉也有可能。
但是,植群它是物種的組合,所以植群的演變講的是:物種的組合在演變,而不是某些種類就一定要消失、一定要換成其他種類。演替是:植群的種類的組合在隨著時間變化。
提問:演替基本概念圖中出現了Softwood Trees、Hardwood Trees。請問什麼是軟木(softwood),什麼是硬木(hardwood)?
裸子植物,針葉樹,叫「softwood」,「softwood」軟木這個字不是來自於裸子植物比較軟,而是來自於木材的構造。當我們把木材切片用顯微鏡觀察,你會看到很多細胞排列在上面。軟木的意思是因為細胞排列得非常整齊,看起來很平坦、細胞大小很一致,排起來很整齊,英文叫smooth,視覺上覺得很整齊、感覺很舒服這樣,所以他們把裸子植物稱為軟木是因為細胞排列很整齊。
相對的,闊葉樹叫「hardwood」硬木,是因為它木材切片的細胞有大有小,大的細胞、小的細胞混在一起,排列不是很規則,看起來覺得很不舒服、很不順,就稱為「hardwood」。
Softwood、hardwood跟質地軟硬沒有關係,因為裸子植物也有硬的,闊葉樹也有軟的,純粹是因為在木材解剖構造上的差異性而產生的一種敘述方式。其實以針葉樹來說,真的是滿多種類的,有的質地軟、有的質地硬。特別是松科的大部分都滿軟的;但是柏科、杉科的,木材就比較硬;至於鐵杉則是密度很高很硬,但是它很脆容易斷。
(示意圖 5. 軟木及硬木結構)
D.先驅與極峰(極盛相)物種
這張演替基本模型圖表示的是:一直隨著時間發展的一個演替的過程,但是當時為了學術名詞的定義,硬是把演替過程切成兩大階段,也就是以紅色虛線切分成前後兩部分。演替的前期稱為「先驅的」(Pioneer)、「先驅的社會」(Pioneer community);當已經發展到後期,也就是成為闊葉樹林的時候,稱為「極相的」(Climax)、「極相的社會」(Climax community),或有人稱為「極盛相」。
硬把演替的進程切成「先驅」與「極相」前後兩階段的結果,導致在一些學派裡就會爭論到底什麼叫「極盛相」,你這一刀到底要切在哪裡?大家的觀點不一樣,引起很多爭論。
所以以現在的發展會認為:所謂的先驅跟極相都是概念,不需要把眼前看到的植群歸類。先驅跟極相的定義是在幫助我們了解演替的概念,實際上我們不會去把植群歸類在先驅或極相,這些名詞只是幫助我們理解而已。
來源:Pidwirny, M. (2006). “Plant Succession”. Fundamentals of Physical Geography, 2nd Edition. 2022/12/15,http://www.physicalgeography.net/fundamentals/9i.html
E.初級與次級演替
「初級演替」與「次級演替」是什麼?在教科書裡的演替基本模型圖裡,把演替劃分成兩類:一類叫「初級」,一類叫「次級」。「初級演替」先被定義,之後才有「次級演替」。
「次級演替」的意思是,比如我們常可以在一個草生地,看到慢慢地會有一些灌木、甚至森林長起來,變成一系列的,就好像那張演替進程圖一樣,但是這個演替是發生在這一塊地方的不同位置。
如照片裡面你看到:這個草地有一些灌木在裡面,這個草地應該慢慢地會被這些灌木所取代,慢慢地被森林所取代,然後演變成後面的樹林,只要時間夠久,最終會變成森林。所以這裡的狀況,我們就把它稱為「次級演替」,為什麼?因為這裡本來就有植物了,已經有植物的時候,再繼續演替下去,這就是「次級演替」的定義。

至於「初級演替」是怎麼定義呢?照片中是火山噴發之後的情況。大家可以預期的是,絕不會有植物活在這邊,因為岩漿是幾千度的高溫,岩漿這樣流過去,不管什麼植物都被燒掉了,所以這裡是光禿禿的。
不久你可能又看到開始有植物長進來,因為附近沒被燒毀的地方的植物又傳播回來,開始長出植物來。植物從一個完全沒有任何植物生命的地方長出來的,叫「初級演替」。

地球上絕大部分的演替都是次級的,以現在來講,初級的不多。就如前面的例子,火山爆發之後,可能是初級演替,還有比如新生成的海島,或一些海埔新生地,慢慢地會有海濱植物進來,那算是初級演替。不過這類的例子畢竟很少,地球大部分的地方都已經有植物存在,當這些地方重新演替的時候,就會形成次級演替。
以上是初級跟次級兩種不同的演替模式。這兩種演替模式在我們森林復育上的操作概念是完全不同的,為什麼?因為次級演替它本來就有植物了,既然已經有植物的話,我們在做復育的植栽的選擇還有作法上,就要考慮本來已經存在著的植物。存在的植物不是只有地上看得到的,還有地下看不到的「未來的植物」。
F.基本模型的問題:時間尺度、干擾、單方向性
教科書裡給的演替的基本模型,問題很大。它有三個問題:第一個是「時間尺度」,第二個是「干擾」,第三個是「單方向性」。以下分別說明演替基本模型的三個問題。
演替基本模型圖裡的第一個問題是「時間尺度」。什麼是「時間尺度」?
我們回顧一下這張演替圖,這張圖說植群會從一年生的草本,發展到最後變成極盛相的闊葉森林,但是變盛之後,再來怎麼了?樹是活的,植群之所以會演替就是因為它是活的,不可能到極相這裡就變成一個終點了,好像拍了一張相片,它就定在那裡再也不改變了。植物還是繼續在活,所以極相這裡的樹很顯然地也會死亡,也會有新的植物再加進來。在這個極相之後呢?就變成一個問號了。所以過去的教科書裡這種簡單的演替模型,完全沒辦法去講「極相之後怎麼了」這件事。
來源:Pidwirny, M. (2006). “Plant Succession”. Fundamentals of Physical Geography, 2nd Edition. 2022/12/15,http://www.physicalgeography.net/fundamentals/9i.html
所以「時間尺度」的意思是,當你把時間看得很遠的時候,這森林究竟會變什麼。這是現在大部分森林生態研究者最想要知道的,因為這牽扯到全球氣候變遷的問題,我們希望去預測一些生態的變化,以及早做一些因應對策。不過這種演替的基本模型是沒辦法解決時間尺度的問題的。
演替基本模型的第二個問題是「干擾」。什麼是「干擾」?其實演替之所以會隨著時間來進程,就是因為有干擾的關係,可是它沒辦法讓你預期發生演替的原因一定是什麼。例如,假設這裡本來是一個湖泊,湖泊慢慢地陸化之後,就有草本植物、變成灌木、漸漸地變成森林,可是假設到灌木這個階段它又形成湖泊了呢?比如來一場很大的水災,或者是地層下陷,這個地方又積水了,它也有可能又變回湖泊。
實際上,在演替過程中間又遇到一些狀況,發生一些變化,然後跑到了另外一條演替的路,這在整個地球的森林演替是常態的事情。實際上你也常會看到,一年生草本之後,樹的小苗馬上就進來了。
所以這個演替圖只是一個理想的模型,它是要建立學習者演替的觀念,可是如果我們只建立這樣的簡化的觀念的話,對演替很多的解讀會是錯的。所以我們做森林復育要考慮演替這個狀況,萬一你只知道到這張基本模型圖,那就很危險了,因為你很可能會做出很多違反森林動態的事情。
再來,這張演替基本模型的第三個問題是,這張圖講說:演替是一個單方向的發展,絕對是從一年生草本這裡變到闊葉林這裡,當然這是一種過度理想化,教科書把它弄得過度簡單。然而實際上到底是怎麼樣的情形,在下一個主題「森林動態」我們會來探討。
提問:如果發生大規模山崩,植被整個脫落,裸露出岩石碎屑,這算是初級演替?還是次級演替?
以我們研究楠梓仙溪的規模來講,楠溪的大崩塌是屬於次級演替,因為它表面的植被被沖掉了,底下的種子庫沒有完全消失,就定義來講是符合次級演替。種子庫裡的種子是活的植物,只是它是以種子的形式存在。
也有可能當這個山崩的規模大到整個地表被切掉很深,比如說1公尺深,露出底下的岩石來,連土壤、種子庫全都消失了,附近也沒什麼植群,這樣就有可能是初級演替,所以要看山崩的規模。
提問:「次生林」跟「次級演替」,這兩個名詞在定義上有相同之處嗎?
「次生林」比較是跟「人的作為」有關。也就是說當一個原始林,(所謂原始林就是人沒有去更動的森林),我們把它破壞之後,或叫作「干擾」,干擾之後這個森林產生一些變化,我們把這個森林稱為「次生林」。次生林是從原始林被干擾之後才形成的東西,所以次生林跟原始林當你用了這個名詞之後,比較會是一個傾向於-有沒有人為介入的問題。
但是演替不是這樣,當然在過去發展裡面都比較考慮自然的這個部分。雖然說「次級演替」也有可能是人為產生的,但是「次級演替」這名詞不會特別強調是人的關係,但是「次生林」是還滿強調人的因素在裡面的。
主題 2. 森林動態(forest dynamics)、更新(recruitment)
我們用右圖來說明「森林動態(forest dynamics)」。基本上還是可以想像成演替基本模型那一條路,但是我們加幾個觀念進來:第一個是,因為我們不知道當演替到極盛相之後,往後會發生什麼事,所以現在的科學研究會傾向的一種觀念是-如果這個植群真的達到一個極盛相的森林,它不會只是停住不動,科學家認為到達極盛相的森林,它會做一個自己的小循環,只要沒有太大的外力在干擾的話,這個小循環會一直在跑這樣。所以科學家們進一步就會想要知道,這個小循環跑一次要多少時間,這個小循環稱為「動態循環」。
為什麼叫「動態循環」?科學家認為極盛相森林應該會在這個地方跑到某個時候,它又會回到原點,所以它是一個循環,又因為它是一直在變動的,所以叫「動態」。
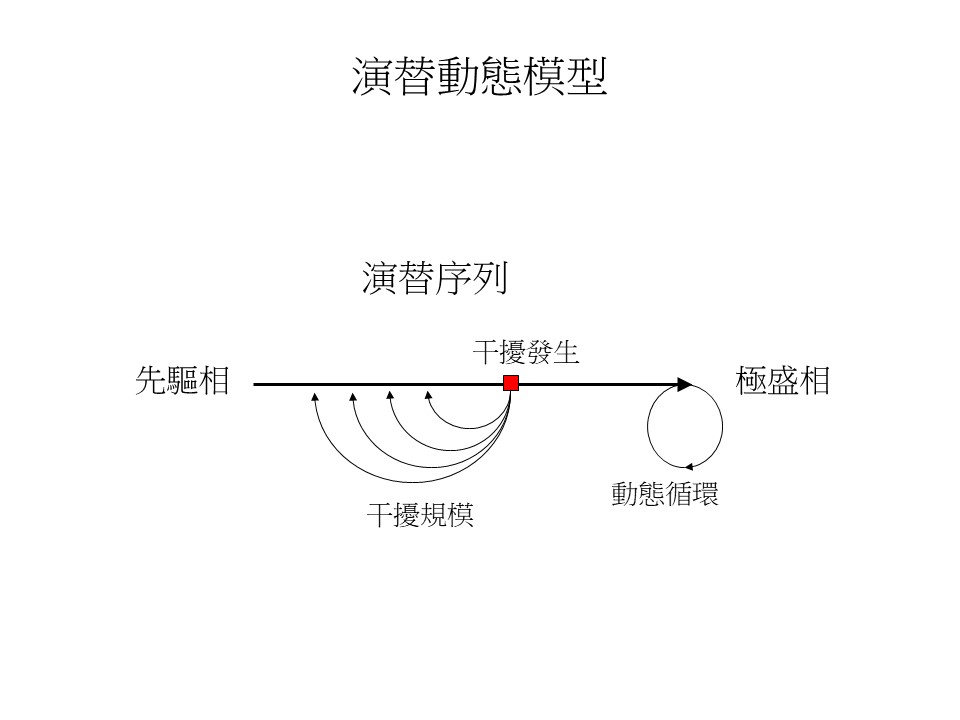
所謂「極盛相森林」是:如果這個森林進入一個穩定的動態循環的話,就可以稱為「極盛相森林」。不像過去看到這森林變成闊葉樹林就說它是極盛相森林。所以在定義上用「動態循環」會更嚴謹一點。
第二個問題是,演替的過程會遇到干擾,遇到干擾怎麼辦?這時候就要看「干擾規模」。比如這個干擾可能只是砍倒一棵樹,這個干擾也有可能是一場大洪水把一片森林都削走了,會有各種不同的狀況。當這個干擾規模小的時候,它可能會跑回去一點點。當這個干擾規模大的時候,它跑回去更多。當然這個模型仍舊是比較單純化,但至少這個演替動態模型把演替基本模型幾個沒辦法解決的問題,把它加進來解釋。
你看臺灣為什麼要設立那麼多動態樣區?原因就是現在在研究演替的人想要知道的演替過程在動態循環、干擾這方面的知識。你看動態樣區幾乎都設在生態保護區,因為生態保護區裡的動態樣區是人為干擾最輕微的地方,所以會假設那邊的極盛相森林會有一個循環,我們希望知道臺灣這些天然林的動態循環要多久的時間,這是有助了解森林的很重要的基礎知識;另外就是在樣區研究這種演替的干擾,當干擾發生的時候森林會怎麼變動,這是要了解森林在改變時所需要的一些知識。現代森林的生態研究大部分就是希望藉由演替動態模型這樣的架構來了解。
森林動態還有一個專有名詞叫「更新」(recruitment)。「動態」其實就是一個森林在「更新」的過程,這兩個名詞是有關連性的。
A.演替的動力來自於干擾
「演替的動力來自於干擾」,為什麼演替會這樣子發生,然後進行?就是因為「干擾」,不斷地有干擾才會進行演替。即使這個森林進入到動態循環了,它還是在循環,它還是在演替。所以在動態循環裡面也有干擾,干擾它會發生在演替動態模型過程裡的任何一個地方,干擾是在演替過程裡面很重要的因子。
「干擾」是什麼呢?就是當我們人類或是大自然對這個森林的作為,讓森林受到一些影響,當這個影響導致森林裡某些植物死亡,植物死亡之後,這森林就會有一些動態的改變。簡單來說,就是某一種外力進來了這個森林,導致裡面的植物死亡,這就通稱為「干擾」。
B.干擾可發生在任何階段
C.干擾發生時間與規模是主要的影響因子
「干擾發生時間與規模是主要的影響因子」意思是說,干擾發生在哪一個時間點,還有干擾的規模大小,是影響這整個演替的關鍵。比如可能是在演替一開始它就發生了干擾,或是在演替中段發生干擾,或者甚至在極盛相的動態循環裡發生了干擾,這些的狀況都會不一樣,它很複雜。
對一個完整的森林來講,它可能在森林的這一區發生一個干擾,另外一區也有發生干擾,所以干擾是不斷地在發生。比如說颱風吹過去,可能是幾棵樹在這邊倒掉了,這邊就開始有一些改變,我們叫做動態。但是可能在一個月後又來一個颱風,在另外那邊又幾棵樹被吹倒了,在這森林裡面不同的地方不斷地在形成干擾,然後就產生了區域的演替的情況。所以干擾的規模、干擾發生的地方、干擾發生的時間等,這些都會導致這個森林動態的不同的改變。
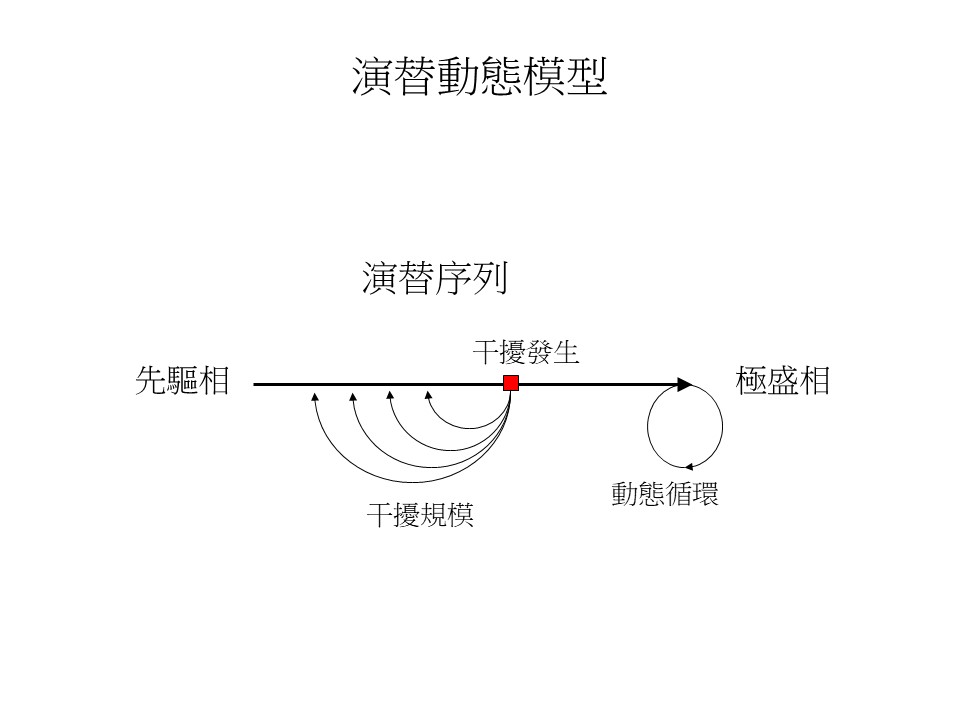
D.干擾若持續存在,不易演替至極盛相植群 >潛在植被
干擾如果持續存在的話,它就不會演替到我們預期會發生的那個植群,在臺灣非常常見這種現象。
以陽明山國家公園裡面的大屯山為例,大屯山最有名的就是賞芒期這段時間,很多台北市民會到大屯山去賞芒。這個地方一大片連續性的草原,是一種叫「白背芒」的植群,這個位置海拔大概是1,000公尺。以蘇鴻傑老師的植群分帶架構的預測,海拔1,000公尺是楠櫧林帶,再加上這個地方雨量很多,它應該是會發展成一個楠櫧林帶的闊葉林,以樟科殼斗科為主。但是實際上這裡整片都芒草,不是楠櫧林,為什麼會這樣子?這就是有干擾持續的存在。

這個持續的干擾是什麼?就是火災。大屯山這地方的芒草的狀況,有點類似大肚山的大黍的狀況,大肚山大黍很多,在乾季枯死變成燃料,容易引發火災。但是大肚山的大黍不斷地發生火燒是人為引起火源的,而大屯山這邊是天然的火災,它不是每年都有火災,它可能十幾年、二十幾年就燒一次,燒完之後植被又歸零,然後又重新長出芒草。闊葉樹在這個狀況下還沒有機會進來,或者它進來不久就被火災燒掉了,闊葉樹只好又重新進來,所以從芒草草原往闊葉森林的演替,就一直被停在芒草草原這邊過不去,這就是持續的干擾。
白茫茫一大片的,不是芒花?!
人家說看芒花哦,可是我常跟人家講說,不是看芒花,是看芒果啦。為什麼?因為芒它開花的時候是紅色的,當它變成白色的時候已經結果了。因為紅色花期時間很短,一下子就授粉、就結果,就變白色的果實了。所以你看到白茫茫的一大片,其實是賞芒果。
E.理想上可以達到一個循環式的平衡,動態平衡,但實際很難
「理想上可以達到一個循環式的平衡,也就是動態平衡,但實際上很難」,這個意思就好像,演替序列發展這條路,會有這個干擾,就一直擋在這裡,當這個演替好不容易跑到這裡來對不對?就又出現了干擾又要退回去,退回去又重新跑,跑到這裡干擾它又來了。所以這個演替序列就永遠跳不到最後面,這樣的話就變成長期是芒草原的狀態。
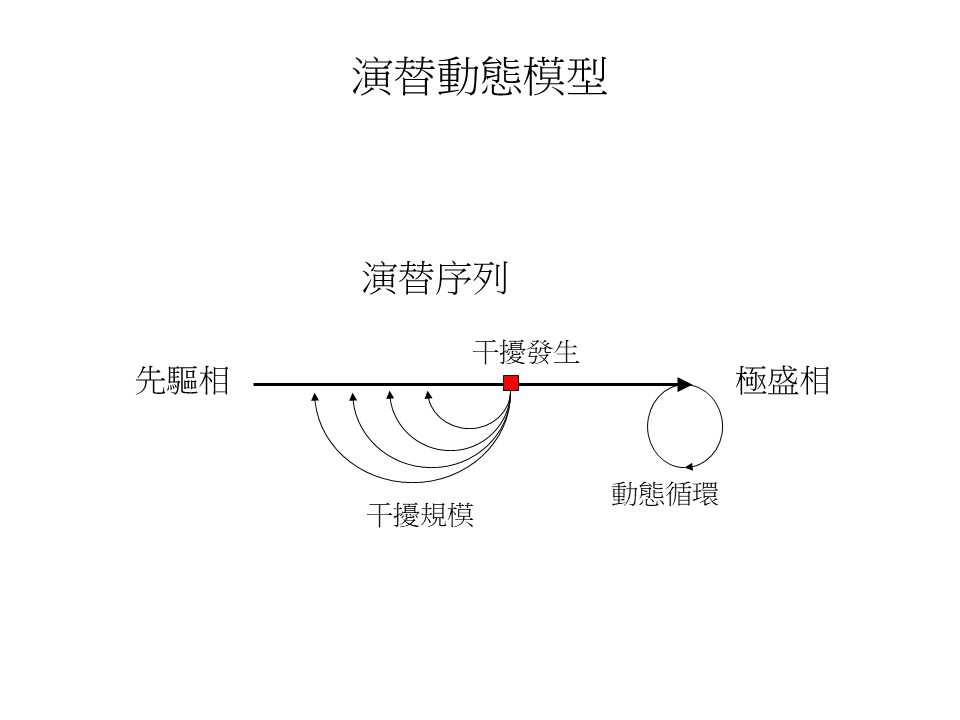
這是大屯山另一個不同的位置,你可以看到它這個地方是有能力變成森林的:這個樹已經長在芒草堆裡面,如果繼續讓它演替,蘇鴻傑老師預測的楠櫧林帶森林就要出現了,但是這裡就是不斷地遇到火災的干擾,演替序列走一走可能就不斷地遇到干擾,又回來起初的點,所以火災的持續性干擾就擋在那邊,潛在森林的演替無法繼續。在臺灣這種情況很常見。

臺灣特別是海拔2,000多公尺以下的草原,幾乎都是持續性的干擾出現的。我們到高山看到玉山箭竹草原是一個非常常態地持續在那邊,但實際上玉山箭竹草原它也有可能是有持續性干擾,不然的話,玉山箭竹草原應該會變成冷杉林,如果照臺灣植被分帶的預測會是這樣子,這個觀念我們叫做「潛在植被」。
演替動態模型的演替序列,最後一個箭號指的極盛相森林,就叫做「潛在植被」。潛在植被的意思是:這個地方最終在極盛相會是什麼森林,但是因為某些干擾的因素這個地方它到不了演替序列的最後,所以潛在植被就是這樣的觀念。
我們在進行的大肚山森林復育某方面是有點推測性質的,我們想要知道大肚山潛在植被是什麼,不然你就沒有辦法完成天然林復育,因為從有紀錄以來的大肚山就是已經是相思樹造林,之後大黍又進來,所以你永遠不知道大肚山是什麼樣的天然森林,這是很困難的一件事。所以我們要根據大肚山的情況來推估它的潛在植被,以潛在植被為藍圖來進行天然林復育的工作。
最後,當演替到達極盛相的時候,理論上那個動態循環是可以持續不斷的循環式的平衡。為什麼會是這樣子呢?我們可以用樹的一生來做說明:當到達極盛相那一邊的時候,森林裡面主要的那些組成樹種,有一些成熟樹它會進入可以繁殖下一代的狀況;這時候它會散播種子,種子出來之後萌發,萌發之後變小苗;然後小苗長大;在小苗長成小樹的過程,這個過程在生態學名詞上稱為「建立」, 「建立」的意思是本來沒有這種樹的地方,小苗進來了,所以這個樹的基礎就建立在那裡了,所以用「建立」這個名詞;再來是,小樹慢慢成長成熟樹。
在這中間的整個過程,不管是種子成熟、小苗、小樹都有可能會死亡,這就是所謂的「動態循環」。樹的死亡在動態循環裡是很重要的部分,因為這個樹如果不死的話,它的數量會暴增,但實際上這不可能發生,所以任何一個階段它都是會有死亡的。
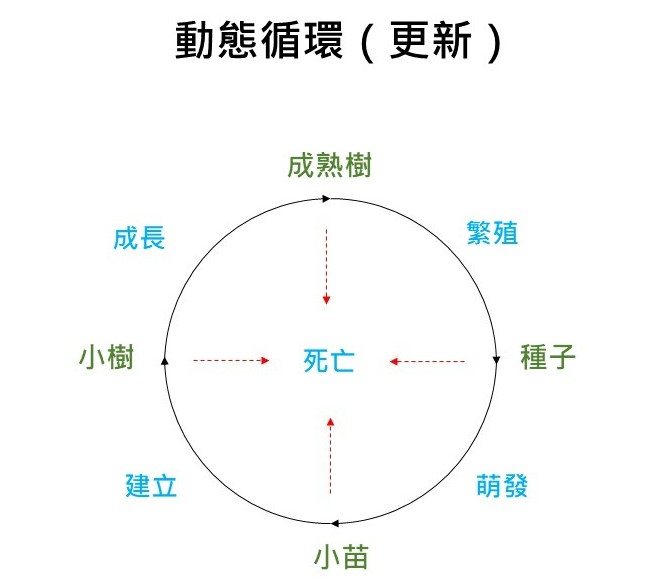
其實有一個最關鍵的位置就是在「小苗建立到小樹」這一段。成熟樹繁殖產生種子,種子量是很大的,種子變成小苗,小苗很多。像協會在做大肚山採種,應該有這種感覺,你真正去知道那個樹的一年的循環之後,你去看特定的時節,在它種子散播出來之後去看,會發現它樹下很多小苗,但是這些小苗如果我們都不要管它,都不要把它採走,或者是設個樣區在那邊觀察,你會發現最後變小樹的,你不要說一棵或者是幾棵,那一年可能一棵都沒有。
所以通常這個瓶頸點是出現在「建立」這個時期,雖然不是每一種樹都這樣,但是以目前我們對樹的了解,「建立」這個時期對樹的成長來講,是一個極為關鍵的階段。
那麼死亡率會有多高呢?我們可以想像一個植物,比如檜木,過去有統計說那種臺灣巨大的檜木一年可以結一百萬顆種子。可是你在那種巨大檜木的附近完全找不到檜木的小苗,完全找不到。它一百萬顆種子全部下去,都是槓龜啊,死亡率很高,很恐怖。
我們有時候回過頭來想,當這個森林裡面的這一棵樹,它只要有它的一棵下一代能存活,它就已經不會滅絕,它的族群數量已經不會減少了。然後,那一棵樹可以活多久?像檜木它可以活一、兩千年,它在這一兩千年裡面只要有1棵後代活下來,在它死的時候取代它,它的族群就不會減少了,對不對?如果它有2棵的話,不得了,如果這森林裡面的族群,每一棵都是有2棵後代活下來,它的族群就2倍了,這很難發生的。所以小苗建立的過程,確實是在生態學一個非常瓶頸的階段。
基本上我們會預期動態循環會這樣子的持續發生,因為樹基本上它會遵循這樣的循環在走。我們把很多很多樹的這種循環總和在一起,就是動態循環。
所以有時候研究動態循環的學者會有兩個做法:一個做法是整個森林做一次整體的研究,比如像我們做動態樣區監測的時候,每一棵都去量;但是也有第二個做法,就是有些人他是針對特定的樹種去了解它整個過程。你有這種大方向的研究,也有這種小方向的研究,把這些不同尺度的研究組合起來,你才能夠比較清楚地知道森林的動態循環的細節是什麼。不過這在研究上真的很困難,需要很大的人力投入。
F.森林動態研究的發展與探討全球變遷議題有密切關聯
森林動態研究的發展跟探討全球變遷議題很有關係,所以現在這個議題這麼受到重視就是因為全球氣候變遷的問題,各國政府他們都想要知道在這種全球化架構下面,一些環境生態、甚至是人類社會的變化,所以森林動態研究的發展就開始受到重視,是跟環境變遷的議題是有關係的。
提問:請問像陽明山白背芒在演替的時間的尺度,如果都不發生火燒的話,白背芒會盤據多久?
在我們舉例的照片裡的大屯山芒草草原,樹就已經進來了,如果在陽明山像照片中這一種等級的紅楠,只要不火燒,大概一、二十年就建立起來。所以我覺得火燒後一兩年內,樹種的小苗就會進來了,因為芒草有一年四季的變化,不是整年都維持這麼茂密,特別在冬天的時候芒草會乾掉,就會有一些空間可以讓樹種的小苗進來。剛好紅楠這一類的種子,差不多在七、八月成熟,種子成熟傳播出來馬上就會萌發,因為紅楠種子還算大顆,剛萌發的種子有自己的養分可以支持,然後到十一、十二月芒草乾掉之後,小苗可以照到陽光,就可以長大,紅楠就有機會進來。像照片裡這些都是紅楠,就是因為它在這樣的時機點,所以紅楠特別多,只要紅楠慢慢起來開始遮陰,樹底下的芒草缺乏陽光就會死掉,芒草一死其他比較耐陰的物種就會進來了。
你看照片的右下方有一株開花中的山櫻,這是鳥類可能在附近吃了山櫻的果實,在這邊上了廁所,這棵山櫻就在這裡發芽。所以在這個芒草草原裡,闊葉樹如果要取代芒草,我覺得是不會太慢,可是最大的問題就是這裡可能不斷有火災干擾,因為那邊的土壤挖下去很多是黑色的、碳化後留下來的植物殘骸。
提問:陽明山那邊的火災是天然的,還是人為的?
陽明山的火災通常是天然的。上一次的大規模火災是2010年,那時我們在做陽明山的資源調查,當時磺嘴山生態保護區發生火災。磺嘴山再前面那一次,1993年有一次大規模的火災,那時我在念大學,我們都很喜歡跑去陽明山看植物,那邊本來有黑松造林,每一棵黑松真的被燒成黑松,樹幹都黑掉了,都燒死了。火災之後,底下芒草就開始長出來,你就看到那種景觀。所以它可能差不多20年會有一次火災,要是這個間隔再拉長的話,闊葉樹可能有機會長起來,一旦闊葉樹起來之後,它就更不容易火災了,因為有闊葉樹之後會比較潮濕、更容易保濕,火災的威力就會越來越小。但是在陽明山,火燒的狀況可能還是會持續存在。
主題 3. 樹冠孔隙 (canopy gap)
森林動態很重要的一個機制就是「干擾」,干擾才能造成森林動態的變化,這是森林研究領域裡很受重視的主題。一般的干擾會形成「樹冠孔隙」(canopy gap),樹冠孔隙是干擾之後一個很重要的產物,會造成森林動態的變化。以下我們來說明樹冠孔隙的幾個重要觀念。
A.先驅相-極盛相樹種 (演變)-> 陽性樹種-陰性樹種 (演變)-> 不耐陰樹種-耐陰樹種
前面提到,在演替的過程裡有一個叫「先驅社會」(Pioneer community),最後是「極盛相社會」(Climax community),或翻譯叫「極相社會」、「極峰社會」,這個是以植群(community)的概念來看的。
我們知道植群是由植物組成的,當我們把這個植物一個一個來檢視的時候,也被做這樣的一個區分。早期分成所謂「先驅相的樹種」(Pioneer species),或是「極盛相的樹種」(Climax species)。在觀念上要把植群跟樹種分開,一個是植群的觀念,有先驅植群跟極盛植群;但是也有對單一樹種的講法,就是先驅樹種跟極盛相樹種。
我們來看一下這些專有名詞的演變:先驅相樹種跟極盛相樹種是早期的名詞,後來不再叫先驅相跟極盛相,就如同前面提過的問題,先驅要多先驅?極盛要多極盛才算?你很難去切分。
所以後來改用一種名詞叫「陽性樹種」對「陰性樹種」。比如大家知道血桐、或構樹,是陽性樹種。久了之後又發現,問題來了,其實這個名詞很奇怪,為什麼呢?基本上所有的植物是喜歡陽光的,沒有光就沒辦法行光合作用,所以學界就出現一種聲音說:所有的樹都是陽性樹種,你說它不是陽性樹種可是它又要陽光,到底要到哪個光度以上是叫陽性樹種,哪個光度以下不是?這又變成一個爭論,所以慢慢地這名詞又演變了。
現今我們用兩個名詞:「不耐陰樹種」跟「耐陰樹種」。
「不耐陰樹種」是指,在比較陰暗的地方,這類樹種就無法生存的;「耐陰樹種」是指,在陰暗的地方,這類樹種可以生存的。這兩類的樹它們都需要陽光,所以新的名詞不再用陽光來區分,而是用陰暗來做區分。用陰暗來區分對樹的生理比較貼切。所以我們盡量不再使用陽性樹種或陰性樹種這類的名詞,以「不耐陰樹種」來取代過去講的陽性樹種。不耐陰樹種這個名詞就更接近樹的生理的真實狀況。
不耐陰樹種在生態學上稱做「r-型的生存策略」。「r-型」是一種定義,「r-型生存策略」的基本條件是:第一個,它會結很多種子。意思是說,這一棵樹的一個繁殖季,會結很多種子;第二個,它的種子是屬於小的;第三個,這種樹是短壽命的,也就是說從發芽、到成長到死亡,經過的時間比較短,所謂的短是指可能一、二十年。如果相對長壽命的樹,比如檜木,它可能可以到一千年,一、二十年對檜木來說當然就很短了;第四個,「r-型生存策略」它的種子傳播距離會很長。通常我們會用以上這四個條件來把樹木歸類,而且這四個條件幾乎都是同時會存在,我們只能說幾乎,當然會有例外。「r-型的生存策略」的樹叫「不耐陰樹種」。
我們可以稍微想想,比如構樹的種子,一般人只有看到構樹上面紅色的球狀的東西,那是構樹的果實,它的種子很小,小小黑黑的。例如榕樹也是,榕類的隱花果裡面有很多種子,實際上榕樹一小顆一小顆是果實,它的果實裡面就是種子,很多、很小這樣。
「耐陰樹種」它的情況就剛好跟「不耐陰樹種」顛倒,耐陰樹種稱為「K-型的生存策略」。耐陰樹種的種子數會比較少,種子比較大顆,樹比較長壽,種子的傳播距離會比較短。

不耐陰樹種的例子,比如「白雞油」。白雞油是臺灣種很多的造林樹種,它的果實是「翅果」,每個果實裡面有一顆種子,可是它一棵樹上面會結很多果實,而且它在很小棵的時候就會開始結果實。
臺灣造林的白雞油,在野外的大樹大概可以到10層樓高,一個樹冠展開都比一間教室還要大。
照片 85. 白雞油
白雞油的果實是一個單片的翅膀,它的種子就在這裡而已,一點點,種子很小,這叫「翅果」。它有翅膀可以隨風散播,傳播距離會比較遠。特別是當它成為一棵大樹又長在山區的陡坡的話,種子可以隨風飛很遠的。
白雞油的壽命不長,我們在野外看到,像林務局的造林可能差不多30年,胸徑都可以到50公分了,長得很大長得很快。這就是比較典型的「r-型的生存策略」,這類的歸類為不耐陰的樹種。
照片 86. 白雞油翅果特寫
這是「山桐子」,在大雪山林道有很多,拍鳥的都很喜歡冬天去看,會有很多鳥在上面吃果實。
這種果實叫「漿果」,山桐子的果實的直徑只有半公分不到,一顆小小的果實裡面就一堆種子。
白雞油、山桐子這類的就是比較典型的「r-型生存策略」,它也是那種 「不耐陰的樹種」。基本上 「r-型生存策略」 跟「不耐陰的樹種」會重疊,可是不太一定。

不耐陰樹種-山桐子
有一年我們幫陽明山國家公園在做山桐子的育苗,就採他們國家公園園區內的山桐子,才採一點點,灑到發芽盤灑了好幾盤。隔年發芽盤密密麻麻地全部都是小苗,我叫學生換盆,他們嚇到了,他們說這個小苗這麼多怎麼換?每一個小小的都是一棵山桐子。我估起來,就一個穴植盤裡面,可能就有1000棵,所以學生他們大概就花了好幾小時才換完一盆的一半,還有好幾盆還沒做,我就說算了,就光這些給陽明山國家公園,應該他們就很高興了。
K-型生存策略 及 r-型生存策略
K-型生存策略及r-型生存策略,這是用縮寫命名。 K-型生存策略的 K是引自德文Kapazitätsgrenze,意思是carrying capacity,乘載量; r-型生存策略 的 r 是引自 maximum reproductive capacity,意思是最大族群的成長率。K-型生存策略及r-型生存策略是指植物的一些生理特性的集合,偏向K型就是K-策略,偏向r型就是r-策略。
不過,我們來討論一些很有趣的、大家比較熟悉案例,例如油桐,油桐到底是不是陽性?以前我們說它是陽性樹種,也就是現在我們講的不耐陰樹種。油桐它是很喜歡在陽光下,但是它的果實裡面的種子很大顆,一顆果實這麼大裡面就一顆種子而已,有時候兩顆,這樣油桐到底是屬於 K-型或r-型哪一種?油桐喜歡陽光,它應該是不耐陰的,依照r-型生存策略理論上它的種子應該要多,可是實際上油桐一棵樹沒結多少果實,它的種子數是少的,種子又大,可是相對來講,油桐其實也算短壽命。所以油桐到底是不是K-型,或者它是r-型?你會發現有很多衝突。
照片 88 油桐果實種子
再舉大家熟知的檜木為例,在林業的估算,大棵檜木一年可以結100萬顆種子,算種子多,而且檜木的種子又很小,可是檜木它是短壽命的嗎?臺灣沒有幾種樹的壽命會比過它,這又該怎麼歸類呢?
所以我們必須要了解一件事情,K-型或r-型這種概念都是我們人為硬把它切出來的,為什麼?因為我們對自然現象要了解的話,就要作「定義」這件事情,先做一些基本的定義,做完這些定義以後,就會發現實際上有很多的例外,再針對這些例外,慢慢地把它們補進來。也就是說,在符合定義裡有一些比較基礎、標準的類型,但是你也一定會看到很多違反定義的狀況。
再舉一個比較常見的耐陰樹種為例:它的果實大,種子大,果實結得不很多,傳播距離也短,這種樹就是「後大埔石櫟」。
後大埔石櫟是靠齧齒類傳播,它的果實成熟時是即將過冬的時候,如果它的果實在樹上沒被松鼠吃掉,掉到地上老鼠不會在現場吃掉而是搬到巢穴去貯藏。如果當年是豐年結果實很多,老鼠來不及吃完,隔年春天有別種的食物長出來了,老鼠也就不吃果實了,它的種子就有機會發芽長出來。因為地面的齧齒類活動範圍通常很小,巢穴就在附近而已,新長出來的小苗也就不太遠。所以殼斗科植物果實的傳播機制讓它的傳播距離很短。
照片 89. 後大埔石櫟
有個經典的例子是在陽明山。為什麼陽明山在森林大破壞之後,回來的主要是紅楠,殼斗科很少?在陽明山上幾乎都沒有殼斗科的,就是在菜公坑山頂也只有看到幾棵殼斗科植物,理由就是因為殼斗科的果實傳播距離非常的短,附近如果沒有殼斗科的母樹的話,這森林只要被大破壞過,殼斗科就很難再傳回來。
可是相對的,紅楠是鳥類傳播,它的種子可以飛過一些地理障礙,從附近山區的森林再傳回來,很快地建立族群在這個陽明山地區,這就是差異。
以上這一節是說明會跟森林更新很有關係的植物的特性,也就是樹種對陽光的需求,會分成耐陰樹種跟不耐陰樹種。
B.徑級結構與樹種的耐陰性
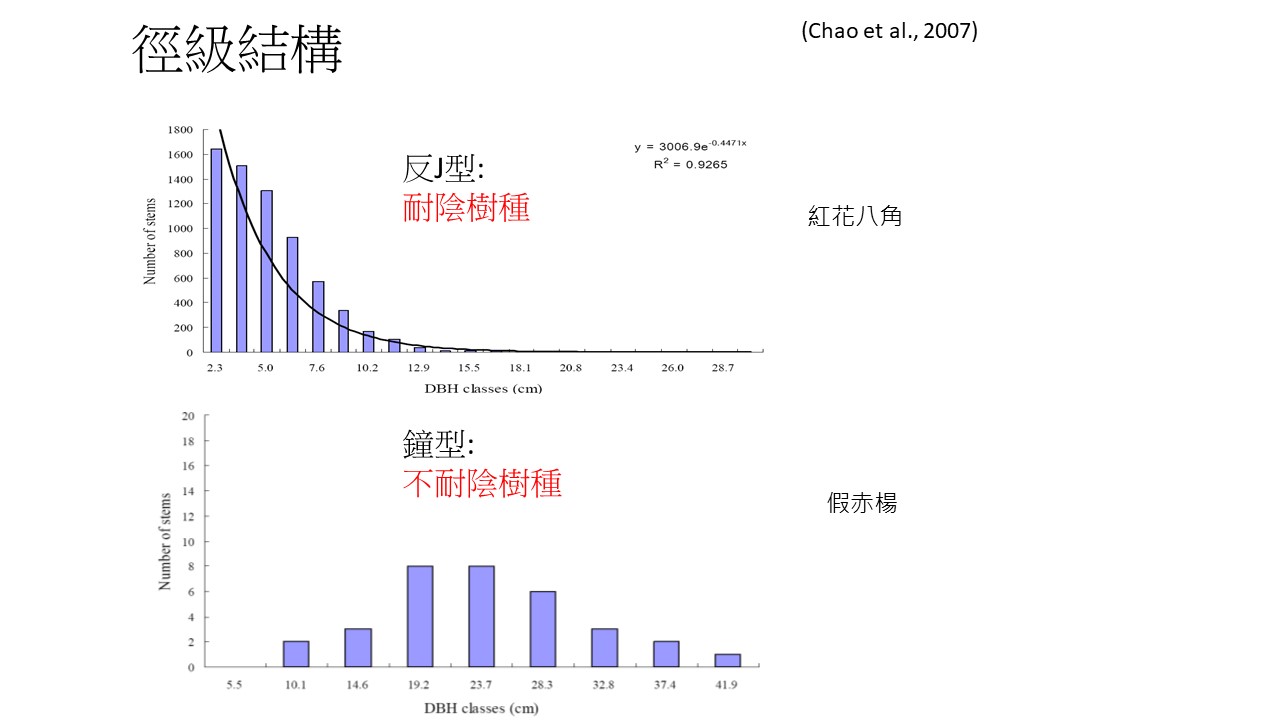
耐陰樹種跟不耐陰樹種另外還有一個很有趣的特性,就是「徑級結構」。徑級結構也會顯露出來這個植物到底是比較偏向耐陰樹種或不耐陰樹種的哪一個。
我們以墾丁南仁山的動態樣區的研究報告為例,那個動態樣區調查到30,000多棵樹,裡面最多的是上方這種「紅花八角」。紅花八角在樣區3公頃的範圍內有大約4、5,000棵。
徑級結構長條圖中,縱軸是株數、數量,橫軸是徑級,也就是胸徑級(DBH classes)。
來源:Chao et al., Species Composition and Structure of the Lowland Subtropical Rainforest at Lanjenchi, Southern Taiwan,
Taiwania, 52(3): 253-269, 2007
「紅花八角」這個植物很有趣,它的徑級結構是「反J型」。「反J型」的意思就是胸徑比較小的,個體的數量比較多。胸徑級越小這個樹就越年輕,年紀越小的數量越多,就代表它這個族群以小孩居多,青壯年就比較少,老年更少,這樣一路下來,就變成反J型的曲線。
只要徑級結構是反J型的,在這個森林裡面通常是耐陰的樹種,為什麼?因為代表在這森林裡它有很多的小苗,所以它通常就是耐陰,因為小苗可以長在那個森林裡面。
「假赤楊」這一種植物的徑級結構是 「 鐘型 」 的。鐘型徑級結構的植物通常是不耐陰植物,年紀小的很少,年紀老的也不多,主要以青壯年居多,形成中間最高、左右比較低的曲線,這代表在這森林裡面,小孩很少,甚至在這森林裡面幾乎沒有它的小苗,但是中間的青壯年的個體很多,它也沒有老年的個體。
「鐘型」徑級結構這種情況是怎麼來的呢?就是在之前的某個時間點,這個植物進來了,在這裡長了很多的小苗,但是隨著這個時間點過去,這些小苗逐漸地長大了。這個時間點過去之後,它也沒有小苗了。所以鐘型顯示的意義就是,在過去的某個時間點它的小苗進來,之後再也沒有小苗進來了;反J型的意義是持續每年都有小苗進來,所以小苗累積最多。
那麼,鐘型結構的小苗進來的這個時間點是什麼呢?這個時間點就是「樹冠孔隙」。在森林的動態裡面我們已經發現,樹冠孔隙的出現使得不耐陰樹種的小苗有機會進來,所以我們可以從主要的、數量最多的植株去鑽年輪,就知道這個孔隙是在什麼時間點發生的。
比如,我們鑽年輪算出這個樹大概30年的樹齡,就是代表30年前這邊曾經有孔隙出現。那麼這個孔隙要多大呢?其實樹種也會跟我們講當時的狀況,因為有些樹種喜歡小孔隙,有些樹種要有足夠大的孔隙才會出現。所以我們就可以知道,在過去,比如說30年前,曾經有一個多大規模的孔隙。
提問:如果在反J型徑級結構出現了孔隙,是耐陰樹種的小苗先上去把孔隙補起來,或者是不耐陰樹種的小苗跟耐陰樹種的小苗兩種相互競爭?
不耐陰樹種通常它的特性是,假設它有機會進來這個孔隙,它的生長速度會很快,很快長高;耐陰樹種的生長速度比較慢,通常是這樣。即使有一些耐陰的小樹已經在,但是耐陰小樹可能才長高一點點,不耐陰的小苗就超越過它了。不耐陰樹種長得真的是很快,比如「山黃麻」就長很快,它的種子很多又很小、壽命短、傳播距離也遠,因為它是鳥類傳播。
但是可能有人會問:如果在反J型出現孔隙的話,不耐陰樹種是比較優勢的,在演替的過程當中,耐陰樹種是不是就機會少很多?其實不會。耐陰樹種它的機會最多,因為它耐陰,所以它一直在那邊等著,它才有機會。就算它長得比人家慢是沒錯,但是它也不會死。它一直等著等著,經過幾次之後,總有一天它就是老大了。總有一天。
講到這個,我就想到世界棒球賽,為什麼日本隊常常在國際賽能夠打到那麼前面?我覺得他們就是這種策略,我覺得日本他就是一個信念,他就是要贏,他就不認輸這樣子,他的策略就是這樣子。所以耐陰樹種就是這樣,它即使長得很慢也沒關係,反正它就死不了,總有一天它就等到了。
C.種子大小通常和休眠能力有關,能夠休眠的種子才能夠進入土壤種子庫
種子大小通常和休眠能力有關。有些植物的種子會休眠,所謂「休眠」就是種子會停止生理活動,不會死掉,等待一個好的機會來發芽,這叫「休眠」。種子的大小會跟休眠能力有關,通常是種子越小越有休眠的能力,能夠休眠的種子它有機會進到土壤種子庫,不休眠的種子就沒這個機會了。土壤種子庫跟森林動態更新很有關係。
D.孔隙是森林動態變化的驅動因子
什麼是「孔隙」?孔隙就是樹冠層開了一個洞。左邊是密閉的樹冠層,你抬頭往上看,雖然也是可以看到藍天,但是你要透過枝葉中間的空隙才看到藍天;但有時候我們會看到像右邊這樣,天空很亮,為什麼?因為這裡可能有一棵樹死了,本來有枝條、有長很多葉子的這個空間,瞬間就沒有障礙了,陽光可以直接照下來,這個狀況就稱為「孔隙」。「孔隙」是現在我們認為在森林裡面最重要的一個機制,如果沒有孔隙這種機制的話,這個森林的平衡就沒有辦法維持。
照片 90. Closed canopy 密閉的樹冠層
照片 90-1. Gap 孔隙
孔隙的出現基本上有不同的規模,主要分成三類:第一類是這棵樹是站著死掉的,上面的枝葉都沒有了,形成一個比較完整、比較圓的這種孔隙出現,這種情況通常也是最小的孔隙,通常是來自於這棵樹自然死亡、或因疾病死亡。

照片 91-1. 主幹碎裂死亡
照片 91-2. 全株傾倒死亡
第二類是主幹碎裂造成的孔隙。有時候你會看到樹它可能是是被雷擊中,樹幹裂開倒下來。為什麼閃電打到樹的時候,樹會裂開?是因為高溫。閃電是很高壓、很強大的電流,電流通過植物導引到地下,又因為植物本身是一個電阻,電流通過植物會產生高熱,把植物裡的水分加熱,加熱速度很快,水分馬上蒸發變成氣體之後,蒸氣就衝開樹幹,所以這個樹是整個被衝開的。樹幹裂開之後,有一部分的樹幹殘留著,有一部分的樹幹倒下來,倒下的過程它會壓死其他的樹,但是它因為不是整個樹都倒下來,所以這種主幹碎裂造成的孔隙,會比第一類主幹直立的空隙要大,但是會比第三類這種全株傾倒造成的孔隙,還要來得小。
第三類的這種,樹木全株傾倒,它是整棵都倒下來,所以傾倒的時候,這個樹幹經過的地方都會被壓倒,這個力量是很大的,雖然像照片當中這樣一棵樹,它的胸徑不是很粗,實際上它倒下來是很不得了的。全株傾倒造成的孔隙,形成的主因是風,在臺灣的山區很常見,因為我們山區的土壤層很薄,當風比較大、又下大雨、地面很潮濕的時候,樹木整個很容易傾倒,它整個沿路倒下來經過的那些樹,全部被它壓垮,形成一個很大的孔隙。
如果你有機會去看一些森林,特別是像大雪山林道、或者楠梓仙溪的林道,那裡有滿多漂亮的闊葉林。當你看到有很多小棵的樹很密集的時候,你注意一下附近,應該就會有一棵大倒木躺在那邊,這就是那棵大樹倒下來之後,產生一個很大規模的孔隙,讓小樹長出來。我們看這個就可以知道,這裡的過去是怎樣,哪個樹倒了,什麼樹進來了,這個其實滿有趣的,你可以做這種調查,就知道那些樹是靠著孔隙這個機制活下來。
森林的動態循環過程
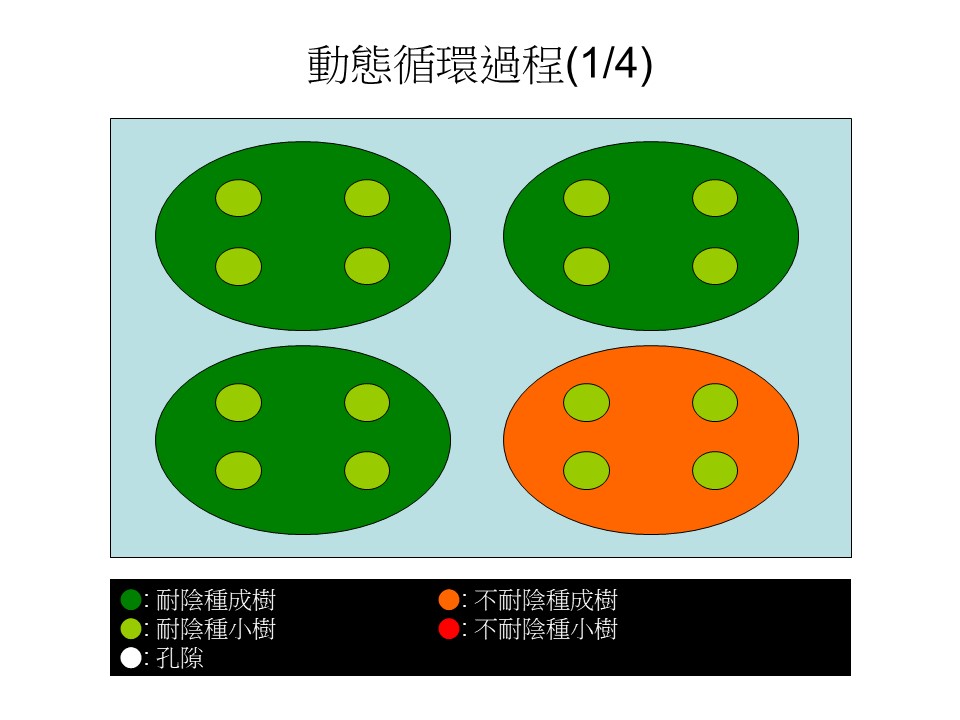
我稍微用四張簡單的圖來說明「動態循環過程」。我們假設一個情況:這裡有4棵大樹,綠色指的是耐陰的樹,深綠色的是大樹樹冠的範圍,淺綠色小圓形是在大樹下方的小樹。
綠色的是大的耐陰樹,耐陰樹底下的小樹一定也是耐陰樹,因為耐陰樹才可以長在樹冠底下光度不太夠的地方,所以耐陰樹的小樹在這裡。那麼這些小樹跟上方的大樹是不是同一種樹?不見得,它們通常是不同種,只是它們都是耐陰樹。
在這張圖裡有3棵耐陰的大樹,底下是它的小樹、或者是別種耐陰樹的小樹。
這張圖裡還有1棵紅色的,是指不耐陰的大樹,不耐陰的大樹底下也有4棵小的耐陰樹。為什麼不耐陰樹底下不是不耐陰樹?因為大樹樹冠下方光度不夠,不耐陰樹無法生長。這是動態循環過程的第一階段。
然後有一個時間點出現了,出現兩個變化:第一個變化是右下方這1棵本來是不耐陰的大樹,因為它是短壽命的,它死了之後,它樹下那4棵耐陰小樹的其中1棵,競爭力比較好的,長起來了,把這一個孔隙補起來。
第二個變化是左上方這棵大的耐陰樹倒下來了,形成一個孔隙。這棵大樹倒下來的時候,壓死了一些耐陰的小樹,可能會有一些沒死、殘留著。假設4棵小樹裡面被壓死了3棵,剩1棵,形成了一個大規模的孔隙。這是動態循環的第二個階段。
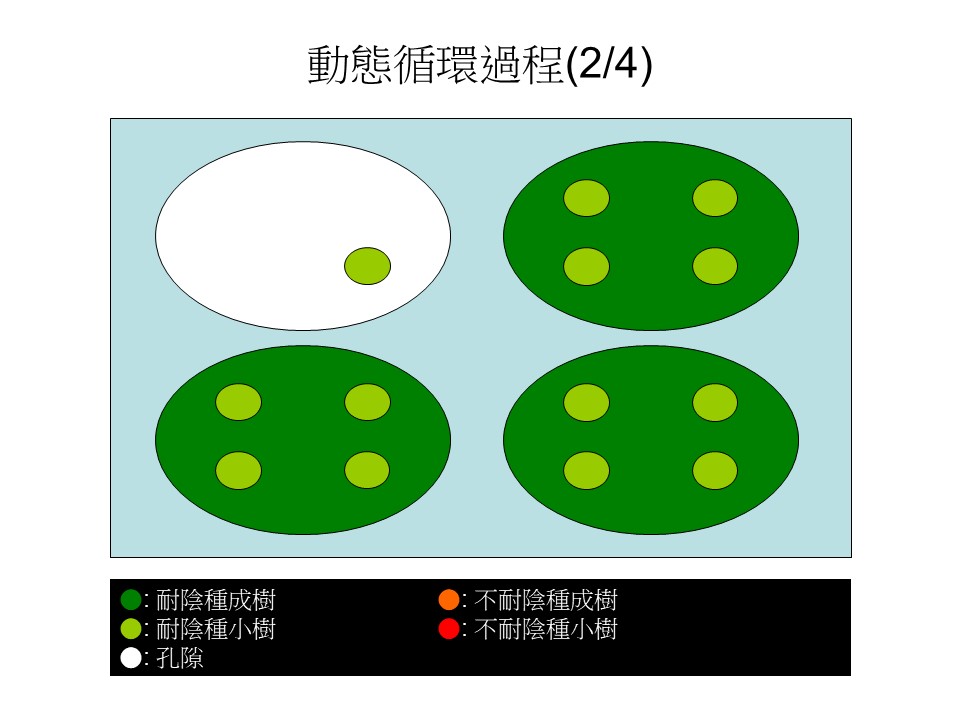
接下來,孔隙裡會出現什麼事情呢?因為陽光很強,直接從孔隙照下來,所以原本大樹底下種子庫裡的不耐陰的種子有機會萌發起來,出現了3棵的不耐陰的小樹(紅色小圓),然後殘留著的一棵耐陰樹的小樹(淺綠色中圓)慢慢長大了。這是第三階段。
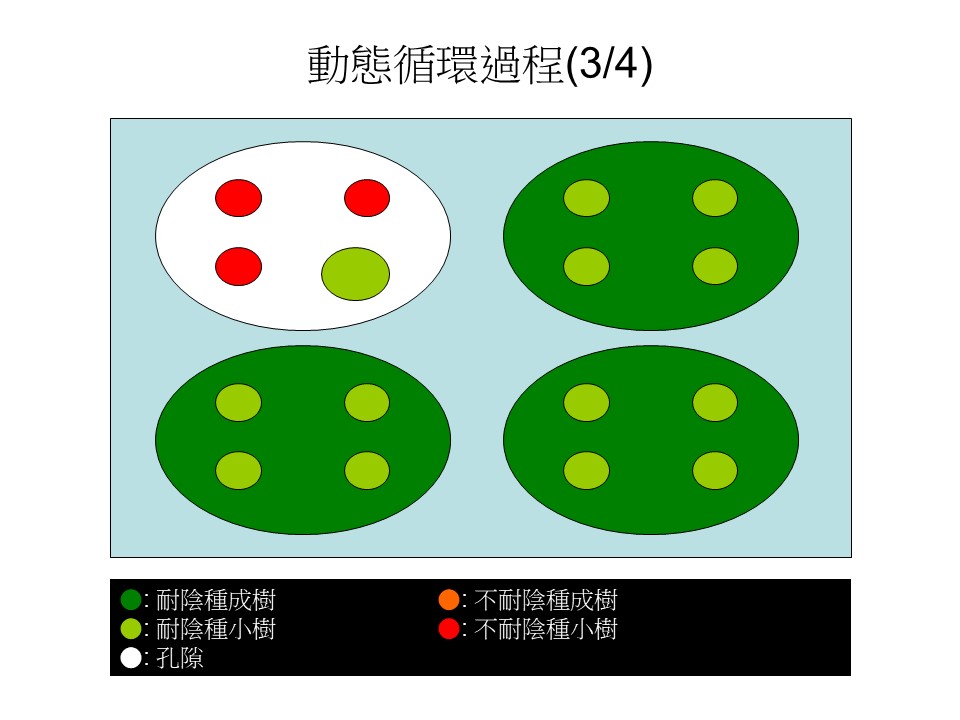
接下來,不耐陰樹種的小樹長得比較快,所以它有1棵可能就長起來了(橘色大圓)佔據了這個地盤,在不耐陰種成樹底下能夠存活的就剩耐陰樹的小苗,其他不耐陰的小樹缺乏陽光,又死了。
可能在這森林的別的地方,又1棵大樹死了又出現大孔隙(白色大圓處),然後孔隙的更新重新又開始了。這是動態循環的第四階段。
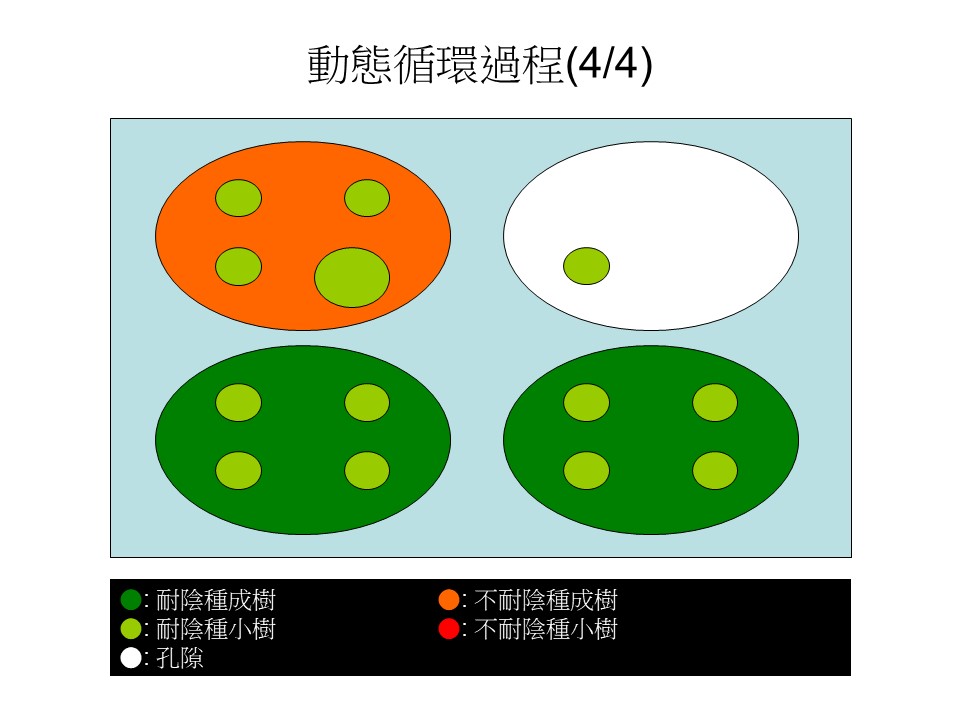
總結來說,森林裡不斷地有大樹死亡形成大孔隙,接著,不耐陰樹的樹種就利用這個孔隙進來了,同時耐陰樹的小樹也在這裡競爭,不久以後可能這些不耐陰樹種就補了這個空缺,但是不耐陰樹種它的壽命短,死了之後又被這些耐陰樹給取代,而別的耐陰樹的大樹可能又會死掉,又會形成孔隙,所以就不斷地在循環。
我們以後在觀察森林動態的時候,就是看這樣的一個動態循環過程,這在臺灣還滿好觀察的,因為我們有滿明顯的孔隙、滿明顯的這些物種。只是在不同的區域,在樹種上可能會有所差異。基本上我們在做這類研究,就是把這些動態循環的每一個細節都把它做出來之後,再拼湊成一個整體。
所以我們可以知道,孔隙的出現很重要,如果這孔隙沒有出現的話,這森林會很單調。最重要的是沒有孔隙的話,那些不耐陰樹種就沒辦法出現在這個森林裡。
過去有些時候我們在森林裡面看到一些陽性樹種(現在稱不耐陰樹種),都會擔心這是不是表示這森林不好?其實不是。這是森林的常態,只要它不是因為人為的關係出現,這其實是常態,而且有這樣的一個機制,耐陰樹種跟不耐陰樹種才可以並存在這個森林裡。並存的好處就是可以增加生物多樣性,這很重要。
像是臺灣山林復育協會進行的天然林調查,其實也可以看一下樣區裡面的鐘型徑級結構的樹,如果你做起來徑級結構反J型、鐘型兩種都有,這就是一個常態。一個正常的森林必然要有比較不耐陰的樹種混在裡面,才是常態。
E.孔隙大小造成不同的干擾規模,進而影響森林動態
孔隙的大小造成不同的干擾規模。大孔隙造成的環境變化會比較大,孔隙存在時間比較久。因為大孔隙比較難癒合,大孔隙底下就會出現比較多不耐陰樹種的小苗。
孔隙大小的一個很大的差異是「大孔隙主要是由新生的小苗把它補充起來的,但是小孔隙並不是由新生小苗補充起來」。
根據研究,小孔隙大部分都是由旁邊的樹擴展過來補充。比如有一棵小樹死了,出現一個很小的孔隙,旁邊的樹於是有了這個機會,它趕快先把枝條往旁邊伸,往小孔隙那邊去搶陽光,所以小孔隙的補充就由旁邊的樹先擴展過去、或是地下莖繁殖先過來補。這一種小孔隙,小苗比較沒有機會來補充。

在自然界的森林裡面,主要的孔隙都是小的。
我們來看一個日本山毛櫸森林的孔隙調查,研究得很清楚。他們調查了一個20.05公頃的山毛櫸森林,觀察了278個孔隙,按照面積分級距,統計孔隙的數量。右圖橫軸是表示孔隙級距,縱軸是孔隙的數量。
研究結果發現,日本這個山毛櫸林裡面,面積越小的孔隙,數量會越多,也就是說,自然界的孔隙其實都是很小的,小孔隙才是常態,大孔隙真的不多。
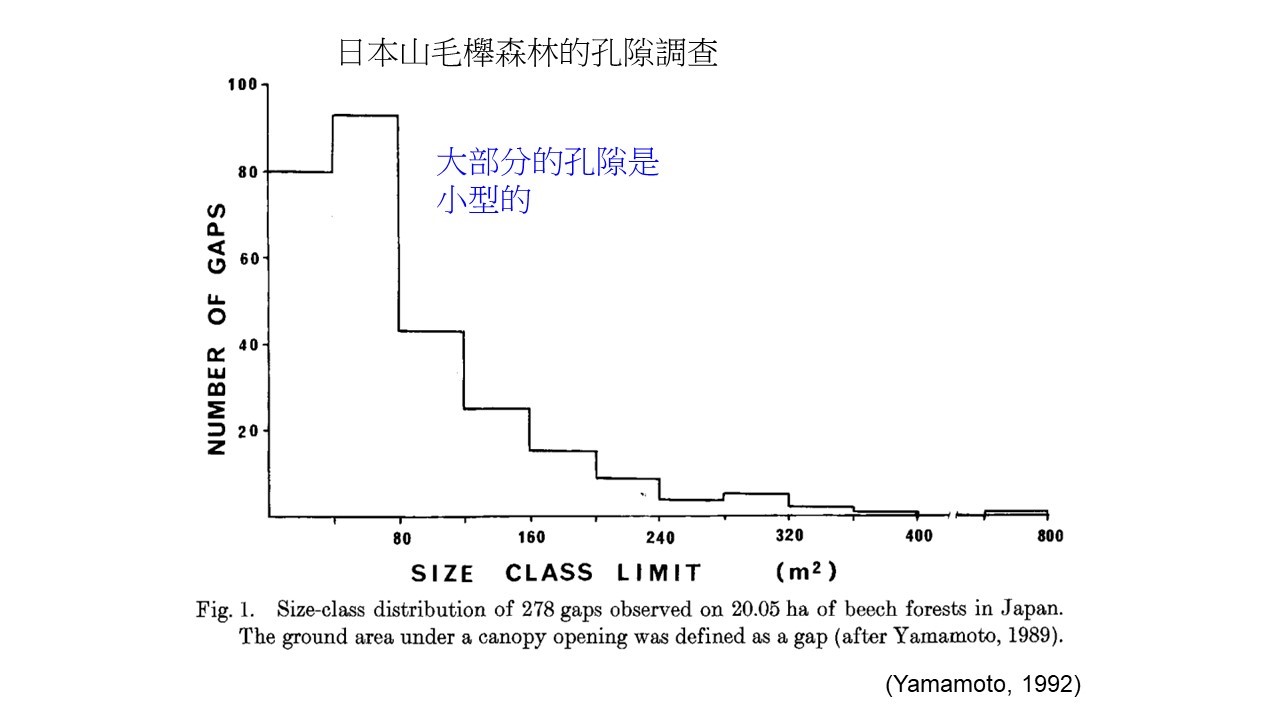
來源:Yamamoto, SI. 1992, Gap characteristics and gap regeneration in primary evergreen broad-leaved forests of western Japan. Bot Mag Tokyo 105, 29–45.
我們舉大家比較熟悉的武陵農場為例,武陵農場的楓樹特別在秋冬的時候,整片都變色很漂亮。「楓樹」一般我們也把它認為是不耐陰樹種。武陵農場還有幾種不耐陰樹種,也是很有名的,就是「栓皮櫟」、「化香樹」,還有「胡桃」。武陵農場這些不耐陰樹種的組合是天然的,雖然楓香有一部份是造林,但大部分像栓皮櫟、化香樹、胡桃等樹種,都是天然的。
為什麼楓香類在武陵農場可以那麼的多?不只是因為武陵農場比較乾燥適合它生長,而是武陵農場那邊的森林有很多小孔隙,這些小孔隙的光度就足夠楓香類的樹種長在那邊了,為什麼?因為那邊的森林幾乎都是落葉樹,落葉樹在夏天葉子很多,沒有孔隙,可是到了冬天葉子落下來之後,形成孔隙了,這時候光可以照下來,所以在從秋天開始落葉到春天還沒長新葉之前,森林裡一大堆孔隙。
栓皮櫟、化香樹、胡桃等的樹種,也是落葉樹,葉子又很漂亮。這些樹為什麼會在武陵農場特別多?就是因為不斷地有季節性的孔隙出現,使它們在那邊能夠存活,這也是另外一種例子。日本的山毛櫸林其實也有點這個味道,只是溫帶森林的物種比較單純,不像我們臺灣這邊生物多樣性這麼高。



F.孔隙提供不耐陰樹種存活的機會->物種豐富度的增加
孔隙提供不耐陰樹種存活的機會,使得物種豐富度增加。孔隙的出現使得不耐陰樹的種子有機會萌發,長成小苗之後跟耐陰的小樹互相競爭,這一點在森林動態循環過程已經有介紹。
G.孔隙扮演維持森林多樣性的重要角色
孔隙扮演維持森林多樣性的重要角色,關於這一個特色我們來看學術研究報告。這是南仁山長期動態樣區的孔隙監測結果,是由屏東科技大學郭耀綸老師的團隊在2003年發表的。
我們看表格中的數字以及長條圖的趨勢,基本上呈現相類似的意義。這個研究是從孔隙發生之後進去做監測,看看孔隙裡面長了什麼樹。
在孔隙發生一個月的時候,原本就存在孔隙下的小苗有26種(109棵)。孔隙發生之後才建立的小苗,一棵都沒有。這個孔隙可能是有一棵大樹倒了。
第20個月,也就是快要2年的時候,發現原本就存在的這些小苗,物種、數量都減少了,可是也發現了一些新建立起來的小苗,有28種(212棵),數量比原來的要多很多。
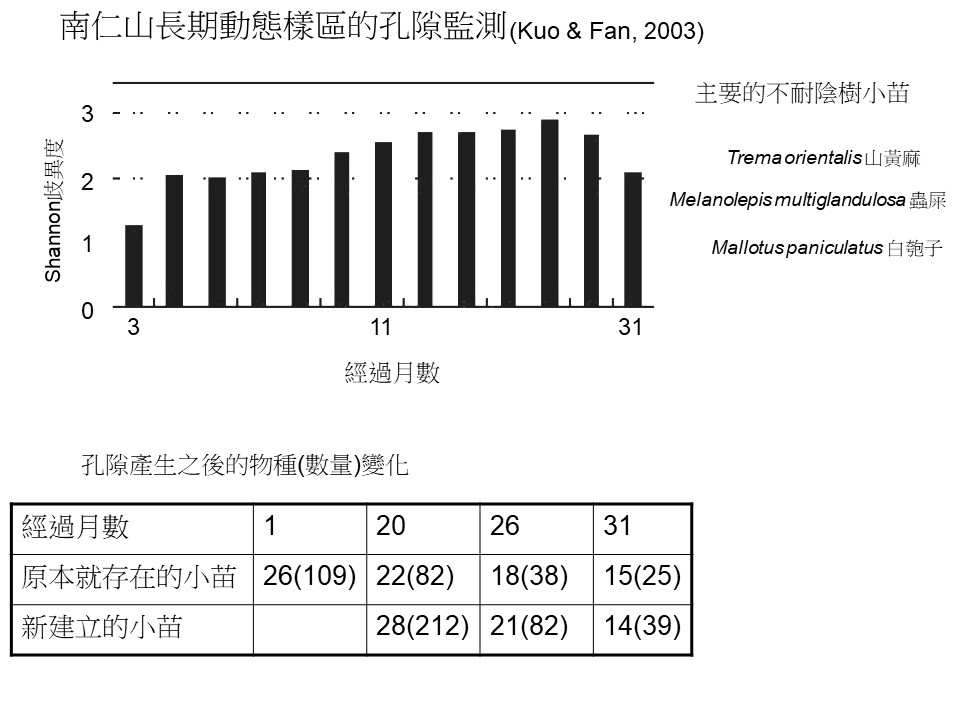
來源:郭耀綸, 范開翔, 南仁山森林倒木孔隙三年間的更新動態,
臺灣林業科學 ; 18卷2期 (2003 / 06 / 01) , P143 – 152
再經過大概半年的時間,第26個月,數量銳減,從212棵變成82棵,為什麼?因為上方的孔隙慢慢被補起來了,底下這個新建立小苗銳減的幅度比較大(21種(82棵)),因為新建立的小苗大部分是不耐陰性的,而原本就存在的小苗通常是耐陰的,所以耐陰小苗減少的幅度比較少(18種(38棵))。
到了第31個月,原本就存在的小苗的物種數,又贏過新建立小苗的物種數了,也就是耐陰的(15種(25棵))贏過不耐陰的(14種(39棵))。
所以繼續再監測下去的話,這些新建立的小苗有可能全軍覆沒,但是上面這一個本來就存在的小苗還會繼續活在這裡,可能還是在持續減少,但是它的死亡率沒有那麼高,等到下一次出現孔隙它就又有機會了。所以一次一次的、孔隙的發生加疊的話,耐陰性的小苗比較有機會能夠來補充。
這個研究所監測到的不耐陰的樹種,突然長出來的、數量最高的前三名有「山黃麻」、「蟲屎」、「白匏子」,這些是在臺灣的次生林很常見的樹種。
在整個森林裡面,白匏子的葉形長得跟其他樹種很不一樣,所以這一種快速生長的植物,它其實有很多不同的生理策略,包括它的葉形的機制、葉面積很大,所以光合作用效率很高。
我懷疑白匏子可能是有固氮菌,因為我們分析過它的氮元素含量,它比陽明山其他的物種還要高兩倍多,在同樣的土地上白匏子的氮素含量特別高,這就表示它吸氮的能力很強。這怎麼來的?可能就是靠著一些跟它共生的微生物達成,像豆科植物有一種能力叫固氮,有點像這樣子。不過我沒有看過任何這方面的報導,我也不是做這種微生物共生的,所以我只能說可能是白匏子有類似固氮的機制。植物不同的生存策略會造就一些不同的生育狀況,它才能有機會活在這邊,不然的話你就看不到它了。
來源:南仁山亞熱帶雨林樹冠層葉片結構之硏究, National Taiwan University Department of Botany, 1993
提問:要怎麼判斷一個樹林是不是到達了極盛相?
這就是過去的學者常在爭論的問題,所以現在的學界已經慢慢拋棄「極盛相」這種想法。在過去如果我們認為這個森林已經是在穩定循環的狀態,就認為它是極盛相。不過現在大家不太用極盛相,比較傾向用「老熟林(old growth forest)」這個名詞來形容這個森林一種很穩定的動態循環的狀態。
怎麼知道森林已經到穩定的動態循環?現在就是用長期監測的方式在做。你如果說10年監測一次這個森林,20年後發現它的變化是差不多的,再10年後,再做一次也是差不多,可能這森林就進入一個穩定的變化狀態,也就是老熟林。最重要的是,你要知道森林永遠一直在變化著。其實我們現在真的比較想要知道的是這森林怎麼去變,然後去推測這森林的未來性。現在主流的意識在這邊。
主題 4. 地被與土壤種子庫
主題4.是「地被與土壤種子庫」。地被與土壤種子庫是在早期的森林動態研究裡比較忽略的部分,因為這兩個研究對象太微小了,一個是長在樹下很小的草本、灌木、或木本大樹的小苗,另一個是躲在土壤裡連綠色的葉子都沒有、看不到的種子。所以過去我們會覺得地被跟土壤種子庫不是很重要,都是研究大樹。但是慢慢地發現,如果不去研究地被跟土壤種子庫,這個問題很大,為什麼?因為我們不知道這些大樹到底怎麼來的,比如有時候我們看大樹的種子掉下來,好像就是沒有長出小苗,但是某個時期小苗又大量出現。於是我們才發現,原來地被,即使是小小的草本,也在影響這個森林。
A.地被是森林更新一個極為關鍵的位置 -> 大型草本植物影響大樹小苗,大樹影響大型草本植物
地被是怎麼去影響森林呢?我們講的「森林的地被」,是指差不多一個人高的高度以下的森林層級。
你下次到森林注意一下,看看長在森林裡不超過一個人高的、地被層的植物,誰的體積比較大、或是覆蓋面積比較大。你會發現面積最大的應該是草本植物,不是木本植物。
也就是說,大型的草本植物它會影響大樹的小苗,因為在那個階段大樹的小苗是一點點在那裡而已,但是草本植物很大叢會覆蓋住這些樹苗,可能會把樹苗蓋到死掉,但也有可能樹苗會躲在這些大型草本植物的空隙裡面,什麼狀況都有可能。慢慢地,等到樹苗長大之後,長上去樹冠層了,樹木會反過頭來影響這些草本植物,所以不同的森林底下的大型草本植物不一樣,是互相的循環,很多研究就指出草本跟木本在森林更新過程相互循環的案例。
特別在臺灣的闊葉林都有很多蕨類,這些蕨類很多體型都很大,過去我們很少去了解這些蕨類對大樹的小苗有什麼作用,影響了什麼?幾乎都沒人在做,因為做大樹的人不認得蕨類,做蕨類的人不認得大樹,過去都是這樣壁壘分明,很少做這種整合的研究,滿可惜的。在國外很多研究都是發現這兩者的關聯性很大。
右表是楠梓仙溪的地被層研究調查結果。楠梓仙溪動態樣區有將近9公頃,我們設了樣帶去做地被植物調查,紀錄小苗,結果發現森林裡有一些冠層樹底下是沒有小苗的。右表清單列出的是在這個動態樣區裡面記錄到大樹的物種有20種,可是在地被完全都沒有這些大樹的小苗。表中特別用紅色字體的是「臺灣赤楊」,為什麼要特別標示它?因為臺灣赤楊在這個動態樣區是一個主要的優勢樹種。在這個動態樣區有四種森林型,其中一種叫「臺灣赤楊型」,這代表臺灣赤楊的數量很多,可是臺灣赤楊一棵小苗都沒有,這很詭異,對不對?

那麼這個森林怎麼更新的?照片中是楠梓仙溪動態樣區的森林,很漂亮。臺灣中海拔因為檜木砍伐的關係,很少有這麼漂亮的闊葉林,雖然楠溪動態樣區也被砍伐過,但因為那邊檜木的數量很少,所以當時是採用擇伐,檜木都砍了,但是大型的闊葉樹沒有動到,才會留下那麼漂亮的森林。

然後,在2009年八月來了一場莫拉克颱風,也就是八八風災,同樣一個溪谷,它的寬度大概100公尺,被土石淹沒,全部都沖掉了。

結果在同年(2009)的11月,在這個森林的地被監測,赤楊小苗就長出來了。所以赤楊的小苗就是這樣來的。
原來在楠梓仙溪溪谷很完整的森林找了半天找不到一棵赤楊小苗,結果八八風災過去之後,小苗就從崩塌地裡面長出來了,為什麼?因為赤楊種子發芽需要光,而且要大面積的光來幫助它發芽。以前這個森林沒有大面積的光讓赤楊發芽,但八八風災過後出現了這樣的條件,所以意味著,楠梓仙溪那一片大赤楊林應該就是這樣的機制形成的,意思是在更之前也曾經有一次這種大規模的山崩,但是在多久以前發生的大規模山崩,可能我們要去鑽年輪才知道。這就是研究地被的重要性。

我用一個簡圖來說明森林更新過程當中,地被層跟樹冠層之間的關係。你會發現地被複雜很多,為什麼?因為這邊的小苗很關鍵,小苗能長大才能夠成為這個未來的森林的補充。
那麼森林小苗要怎麼長大?小苗會面臨很多競爭,比如被動物吃掉,像在楠梓溪仙,動物影響就很嚴重,因為那邊食物很少,所以連很難吃的十大功勞,也被動物吃得一乾二淨;再來是疾病,小苗可能自己就有一些真菌、細菌、或是病毒等這些感染死掉;再來是要跟草本植物的競爭;再來可能是會天然的死亡,比如基因比較衰弱之類的。假設這些沒有被吃掉、沒有病死、沒有被競爭、或沒有自己死掉的小苗,它繼續有機會成長的話,才會變成大樹。所以小苗是很關鍵的。
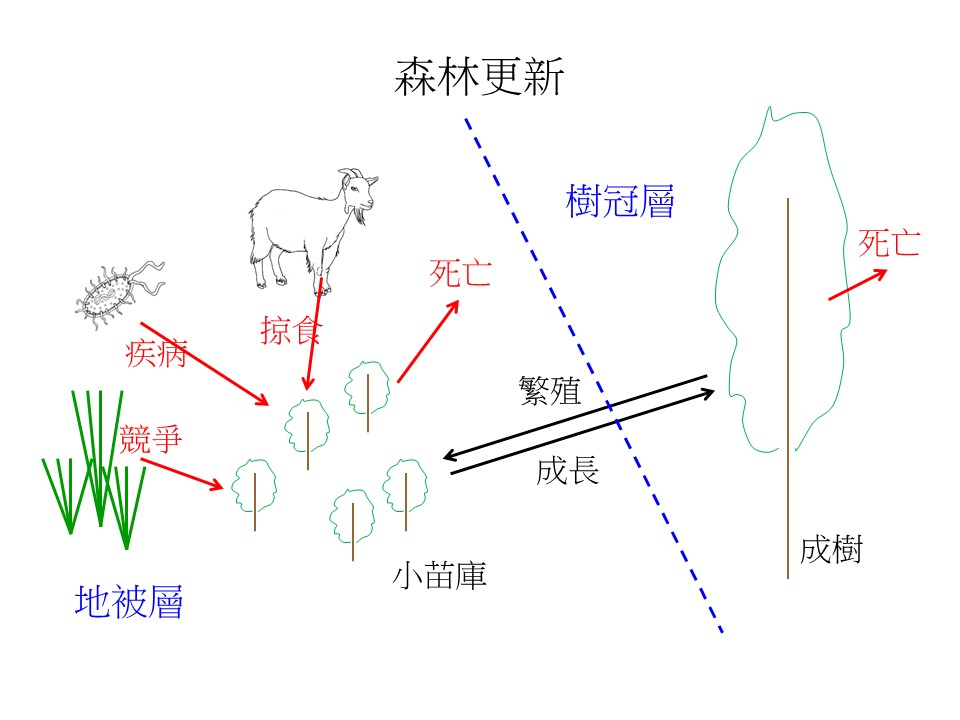
我們通常在森林裡面看到的小苗也滿有趣的,它通常會形成「小苗庫」的形式。什麼叫「小苗庫」?就是當你看到一棵小苗,通常它旁邊應該有很多棵跟它同種的小苗,很少看到單一棵的。當然也有很多生態學假說來解釋這種現象,但是我們看到是這個情況。這小苗庫怎麼來的?有一個很關鍵的就是「土壤種子庫」。
我們先來看一下,在森林更新的過程,草本植物怎麼去影響這個競爭。
這是日本的山毛櫸森林。山毛櫸森林底下密密麻麻都是箭竹,只要有箭竹在的話,山毛櫸完全都沒有小苗,因為山毛櫸種子落到地面,箭竹下面的光度非常低,山毛櫸種子完全沒辦法發芽。
山毛櫸沒辦法發芽它要靠什麼?靠箭竹大規模開花死亡,死亡之後你就會看到很多山毛櫸的小苗就出現。

過去臺灣做山毛櫸的研究也是類似的狀況。他們發現臺灣的山毛櫸好像都只有大樹、沒有小苗,結果發現是箭竹的關係,所以後來就有比如移除箭竹這類的做法,讓山毛櫸有機會發芽。
還有一年,國家山毛櫸森林步道森林下的箭竹大規模開花死亡,宜蘭大學有去做監測,發現山毛櫸在那時候就大量冒芽出來,它就是靠這個機會。這就是地被去影響小苗很有名的例子。

我們接著來說明,小苗庫怎麼出現的?小苗庫的來源就是「土壤種子庫」。
B.土壤種子庫往往不是直接補充耐陰樹種,而是加快不耐陰樹種在孔隙建立的速度 (注意多數植物一年只有一次果熟期)
這是在臺灣南部多納做的一個研究報告,結果顯示在多納林道的森林種子庫,調查到有23種植物種子留在土壤裡面,其中有22種是先驅種,也就是不耐陰的樹種。非先驅種,也就是耐陰的樹種,只有1種。換句話說,不耐陰的樹種要怎麼樣在這個森林裡面很快地就建立起來?

來源:台灣林業科學19 (1): 33-42. (2004)
https://www.tfri.gov.tw/News_Content2.aspx?n=7498&s=10421
過去我們有一種觀念,就是當孔隙形成的時候,森林底下可以照光了,我們就會講「因為不耐陰樹種是r-型策略,種子傳播的距離很長,很快地就傳播到這裡,就長出來了」,以前大部分都是這樣做解釋。可是這個孔隙再怎麼大也大不到哪裡去,這個種子怎麼那麼厲害就剛好可以傳播到這裡?後來有更多的研究之後,我們發現過去那種解釋有問題。
八八風災後出現赤楊小苗的例子跟我們講一個很清楚的事情:赤楊的小苗是土壤種子庫來的。為什麼這樣講?因為通常來講,臺灣赤楊的花期是10至11月,果期是11至2月,落葉期是11至12月。那麼你想想看,2009年八月八號風災的時候大規模的孔隙才形成,經過短短3個月,在11月的時候赤楊的小苗已經在這邊發芽了,這中間如果沒有任何赤楊的種子成熟的話,根本沒有機會來,你不要說傳播,赤楊在11月正開始要結果,它還沒有種子。
所以,很多不耐陰樹種為什麼可以很快地建立在這個孔隙上,是因為它老早就在這個森林的土壤裡面等了好久了。這個等待的時間可以多久呢?國外以前他們對某些物種的監測結果是這個等待期可以超過一百年,這個種子在土裡面等了一百年,他們把種子挖出來拿去發芽,種子還活著、長出小苗這樣子。所以我們現在對孔隙的更新有一種新的看法就是,孔隙的更新主要來自於土壤種子庫。
所以不耐陰樹種可以那麼快進來孔隙,是因為它們大部分都是屬於小型種子,它們的種子可以進入到土壤種子庫,而耐陰樹種就不是這樣。耐陰樹種子比較大顆,沒辦法休眠,所以耐陰樹的種子通常當年度如果不發芽、或發芽完沒有好條件給它成長,它就死掉了,所以耐陰樹是一年一年要補充種子,這是它更新的速度慢的主要原因。
對土壤種子庫的瞭解很重要,但是我們臺灣這一類的研究真的少。我們過去在楠溪曾經有做一次這種調查,做這個真的很困難,包括認小苗、去採樣、或者做一些發芽的操作,這些都有難度而且很耗人力,所以這在臺灣過去做很少,我看過的報告沒有超過十個。
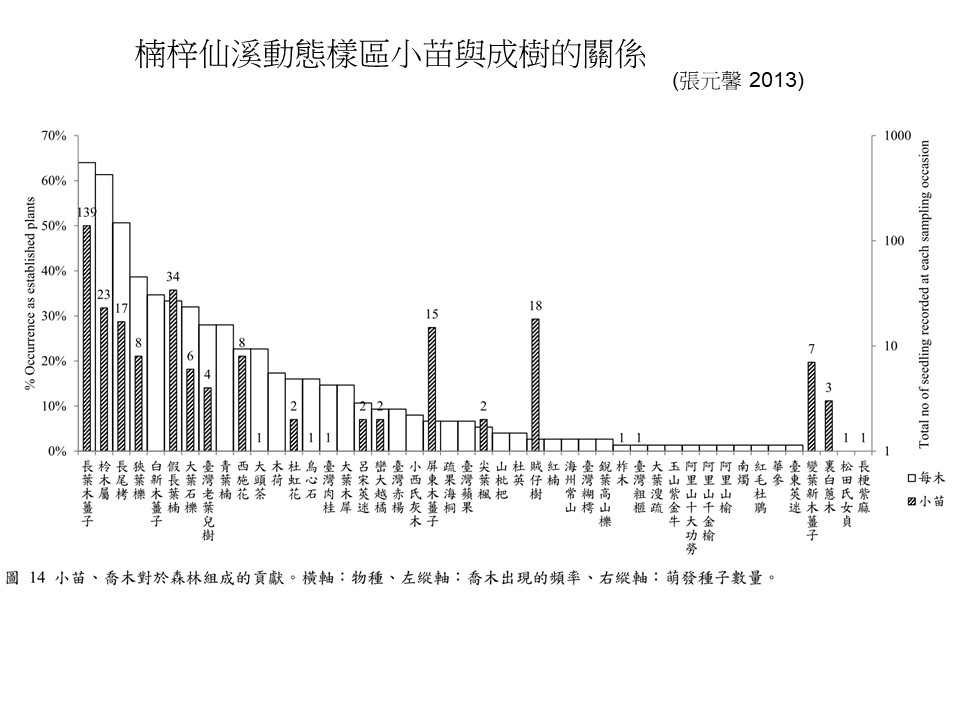
我們舉例楠溪的動態樣區來說明小苗與成樹的關係。每木(成樹)是白色的長條圖,小苗是斜線的長條圖。橫軸是物種名稱,分別表示不同的物種,左縱軸是成樹(喬木)出現的頻率,右縱軸是萌發種子數量。
從圖中可以看到,如果就楠梓仙溪動態樣區的大樹跟小苗來看,它們兩個關係基本上是類似的,也就是說,如果大樹數量很多的話,通常它的小苗也很多,這個趨勢是一致的。但是當然你會看到一些例外,比如有一些大樹很少可是它的小苗特別多的、或是有大樹但是幾乎沒有小苗、甚至沒有小苗的,但是大部分你看到這兩個長條圖的高度的趨勢是一致的,這代表小苗跟大樹的關係你很容易去解釋。可是土壤種子庫跟大樹的關係就不是這樣。
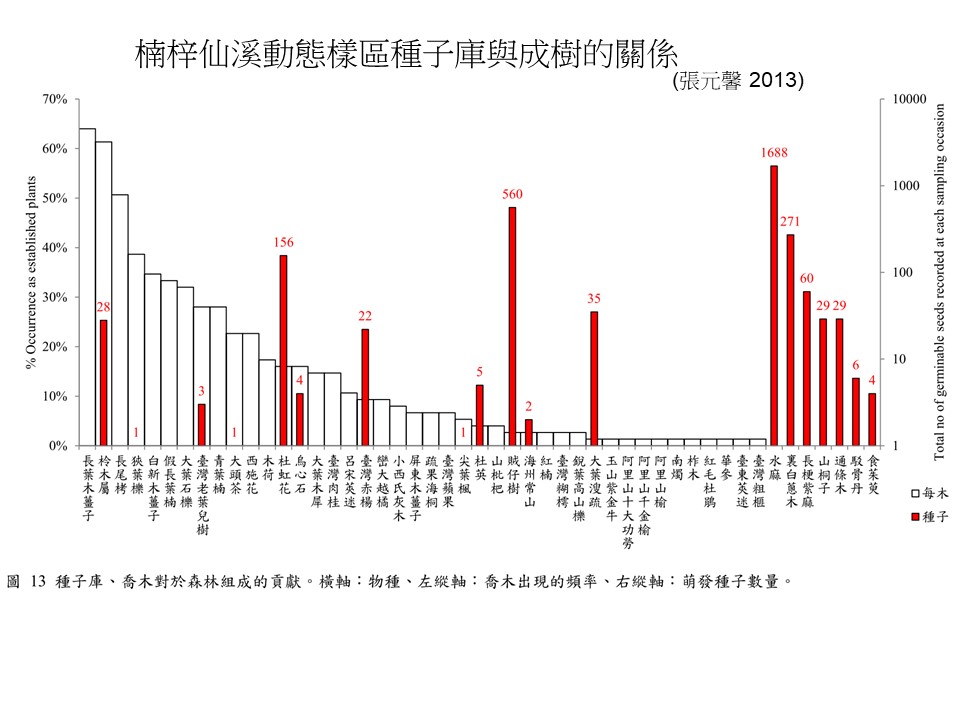
來源:張元馨, 2013, 楠梓仙溪森林土壤種子庫組成之研究, 臺灣師範大學生命科學研究所.
這張圖是種子庫與成樹的關係。土壤種子庫跟大樹的關係非常非常的薄弱,大部分的大樹幾乎都沒有任何種子在土壤種子庫;少部分的大樹在土壤種子庫種子很多;甚至有一些大樹根本不存在這個森林,在土壤種子庫裡有很多很多它的種子。
你看看最主要的這些樹種,幾乎在土壤種子庫都找不到它的種子。另外這些不耐陰性樹種的種子,它在等森林裡孔隙的機會。
所以耐陰樹種跟不耐陰樹種這兩個是很不一樣的:在一個比較好的森林裡面,耐陰樹種以小苗的形式在大樹下面等著,不耐陰樹種以種子的形式在土壤種子庫等著,大家都在等機會,都在等機會。
來源:張元馨, 2013, 楠梓仙溪森林土壤種子庫組成之研究, 臺灣師範大學生命科學研究所.
主題 5. 森林動態觀念在森林復育的疑問
本章的最後一個主題是,森林動態觀念要怎麼應用在森林復育。這裡提出兩個議題來探討:第一個議題是,是否需要根據演替序列進行植栽?第二個議題是,如何達成樹種多樣性?
A.是否需要根據演替序列進行植栽?
蘇夢淮老師:最早蔡理事長在想說,我們在做大肚山森林復育要照演替序列來做,就是先種不耐陰樹種,然後種耐陰性樹種,照這樣的進程來進行。我那時候的想法是,「我們不需要去種不耐陰樹種,直接種耐陰樹種就好了」,當時我的回應是這樣子。
那麼我們來討論,大家覺得我們在做森林復育到底要不要照著演替序列、遵循自然的這一個路線來種,也就是先種不耐陰樹種,一段時間之後,我們再進去種耐陰樹種,種在不耐陰樹種的下面?有沒有必要性?
蔡智豪理事長:當時跟蘇老師討論是說,大肚山就算我們不去種不耐陰,本來整個山坡就都是不耐陰樹種了,那麼到底要種、還不種?其實大肚山不耐陰樹種沒辦法建立起來,主要是頻繁的火燒的干擾。但是如果我們在操作上來講,比如舉例台中都會公園不會有火燒,那我們需要先從白匏子、或構樹開始種嗎?還是我們就直接朝耐陰樹種開始種?然後也發現說,其實耐陰樹種不是有陽光就全部長不好,還是有一些耐陰樹種在陽光下也是可以慢慢起來。當時在討論的時候是,耐陰樹種可以跟不耐陰樹種一起長的,我們在大肚山的這些保安林的荒地上,我們補植耐陰樹種進去之後,耐陰樹種跟不耐陰樹種它們可以在同一個時間慢慢長起來,這是當時討論時的想法。
蘇夢淮老師:我覺得這是開放性問題,大家可以來思考。畢竟未來如果要操作的話,我們在森林復育面臨的第一個問題就是:到底要選什麼樹苗?除了我們要選在地樹種之外,現在大家慢慢有更專業的知識之後,就要去考慮這些樹種的一些生理特性,比如耐陰或不耐陰,可能在森林動態裡面,這些樹種各有各自的特色、各自的重要性或地位,這樣到底要不要兩個都去種,或是要先去種不耐陰樹種?
學員A:我想到一個面向是,耐陰樹種要在林下慢慢長,如果它不是在林下長,它會有什麼差別?因為樹都需要日照,它的光合作用的型態可能是不同,所以耐陰樹它才會是在樹下,它接受的日照少一點,但它慢慢會長大;如果耐陰樹它在一個日照強的地方,如果是以農的角度來看,這個耐陰樹會不會是比較怕?比較怕的意思就是,長得快不等於長得好,我在想會不會像類似這樣?
學員B:如果是耐陰樹種的話,我們在都會公園是在開闊地面,從小苗、種子開始種,那是全日照的環境,這樣耐陰樹種在全日照底下會長得好嗎?這是我的疑問。
蔡智豪理事長:就我們的現況來看,一開始種耐陰樹種小苗,它是長得慢,它不一定長得好與不好。如果我們跟不耐陰樹種來比較的話,不耐陰樹當然長得比較快,但是耐陰樹不見得在全陽光下會死掉,它只是長得比較慢,但是當它到達一定大小的時候,它就不怕陽光了,就很特別,耐陰樹種只要超過一定的大小之後它就不怕陽光,它成長的速度就不會輸給不耐陰樹種,也就是耐陰樹種它必須要大到一定的程度它就會往上衝了。我覺得是如果耐陰樹種從小苗到不怕陽光、可以往上衝的那個階段是我們可以忍受的階段,可能只有三年、五年,它一過了那個瓶頸它就衝上去了。
蘇夢淮老師:我補充說明,其實就對同一種樹來講,成樹階段跟小苗階段的生理需求可能是不一樣的,這種例子算是很常見。耐陰性樹種確實一開始就讓它接觸強陽光的話,它不會表現得很好,可是當它長大到某個程度,它就很喜歡陽光。耐陰性樹種它就是這森林裡面,我們認為未來會長成最大的樹的那一批,你去看它那個大樹一定是直接接觸陽光,在正常的森林是這樣子。所以耐陰樹種的成樹一定不怕陽光,可能怕陽光的階段就是在它小的時候。
為什麼耐陰樹種小時候會有這個特性,長大不會有?當然這要靠很仔細的細部研究去解答的問題,但是就現狀來看的話,你會看到耐陰樹確實有點像符合它自己本身在森林的一個地位:它小時候喜歡躲在樹下,但是它在等那機會,等到它夠大的時候,等到機會來的時候,它就突然長很快,會有這個特性。
這也許就是未來我們在做森林復育一個很好的切入點,就是一開始我們可能兩者一起種,我覺得不用有種植的先後順序是因為你兩者同時種下去,耐陰樹種它長比較慢,不耐陰樹種它一下子就會長很快,就有一些樹林,那個樹陰自然就營造出來了。
另外一個就是,我們在不耐陰樹種的種植可以採取種子庫的思維。種子庫的思維就比如說,在大肚山我們也許可以嘗試一種作法,把地被清除,觀察看看會長出什麼植物,了解種子庫的狀況,可能過去也沒有人做過這種試驗。大肚山經常火燒,火燒能不能算是地被清除?也許也可以算,但是不曉得大肚山的土壤種子庫的深度到多深,如果土壤種子庫比較淺層的話,種子還是會被火燒死。
如果我們用人工去清除大黍、或者是清除一些比較密的灌叢,讓它維持一段沒有大黍的時間,大黍一定會再冒出來,一冒出來我們就馬上把它除掉,看看大肚山這裡天然的狀況會長出什麼不耐陰樹種。有這樣的基礎資料的時候,我們在做天然林復育就比較好操作。
我現在假設,比如構樹或血桐會在這土壤種子庫裡面出來的話,我們就不太需要特意花時間、花心力去種它們。我們就是在那個地方整地過之後,讓構樹或血桐自己長起來,然後把耐陰樹種種進來。有時候你去看不耐陰樹種,它雖然在野外長很多,可是不太好種。在這個狀況之下,你倒不如讓大自然去種它,而像耐陰樹種這種比較不容易在大自然存活下來的,我們再用人力去幫忙。這樣的話我們可以省很多力,而且可以符合演替的法則。
觀念:為什麼可以在大肚山野外操作種子庫萌發法?
過去我們做這些土壤種子庫的實驗,我們不是去挖土回來把種子一顆一顆從土壤裡面挑出來,挑不出來的,因為不耐陰樹都是很小的種子,幾乎跟沙子一樣。所以我們都是用萌發法讓它發芽,怎麼讓它發芽?你把土挖出來之後讓它接觸全陽光,它就發芽了,不耐陰樹種子就是在等這個光,全陽光對它來講就是孔隙的訊號,所以我們把它挖離森林,帶回苗圃或溫室,給它足夠的陽光,它就長出來給你看。所以我覺得我們可以在大肚山直接在野外操作。
蔡智豪理事長:大肚山經常會發生火燒,為了防火,大黍應該要經常清除,可是保安林的防火帶有水土保持的問題,林務局規定防火帶上清除掉大黍一定要種樹,不種的話會被認為是山坡地裸露,一定要種樹。這樣的話,可以種在防火帶防止跳火的原生植物有那些?
蘇夢淮老師:我覺得有一個做法就是把這邊的森林物種都種看看,反正你預期短期可能沒辦法解決這問題,到時候用這機會做一些實驗,看這邊原生的樹種裡面哪一種效果最好。可以種「木荷」,木荷是林務局最常用的防火樹種,你可以看有沒有可能從附近找種源。木荷用盤子萌發還滿簡單,它很會長,它很耐陰,小苗也可以在森林裡面。
蔡智豪理事長:我再釐清一下,以大肚山的現況來講,許多木本植物火燒後都還是活著,可以再萌蘗、再發芽。現在我們在處理的防火,與樹木有沒有被燒死沒有關係。
蘇夢淮老師:最重要就是讓火不要蔓延,防火樹種的功用是在讓火不要蔓延。大黍清除之後,種什麼樹對於防火是有關係,像有些植物的樹冠比較沒那麼密實,底下光度會比較強,大黍還是可以長,可是有一些樹冠比較密的,底下不會有大黍,像是「榕樹」的樹冠很密而且它擴展很大,樹底下應該是會很乾淨沒有長草,榕樹也可以值得一試。不過,「雀榕」會落葉,一落葉可以透光,就會提供大黍生長的機會了,雀榕就不是一個好的選擇。可以種一般的正榕,正榕不會落葉,正榕在大肚山也很多。
其他的原生樹種的話,「構樹」的樹冠層很薄,從葉間孔隙下來的光度比較強,大黍還是有機會。如果要種構樹的話,要種得很密,另外要混搭著其他樹來種,要補滿孔隙。
大肚山發生火燒的問題,大黍就有點像是炸彈的引信在那邊,所以要把引信剪斷,把大黍先剪斷、把大黍除去。所以我覺得應該就是要盡量種樹冠大的,榕樹可以試,榕樹的汁液還滿多的,也不容易著火。再看看大肚山還有哪些常綠性的植物,大肚山如果有「水同木」的話,也可以嘗試。
大肚山目前的環境條件對水同木來講,可能會比較乾燥,但是如果你讓水同木長起來,那個環境就會慢慢被水同木轉化了。其實演替也是這樣子。為什麼這個樹會進來這個地方?因為這地方有這個樹的合適性,但是這個樹進來這個地方之後,它也會改變那個環境。比如水同木如果進來的話,慢慢地環境會變潮濕,降低火燒的機會。剛開始你要種水同木也許困難度會比較高,要多一點人力去照顧,直到它建立。
以上這些討論純粹從防火帶的功能來考量,不是從天然林的角度。關於防火有很多操作的方式都可以再討論。
B.如何達成樹種多樣性?
森林動態觀念要怎麼應用在森林復育,我們要探討的第二個議題是,如何達成樹種多樣性?
過去都有所謂的目標物種,比如種很多殼斗科、或樟科在那個地方,這代表未來那個地方可能是最極盛相的森林。可是在我的看法,我們應該適度保留一些機會讓先驅樹種在裡面生長,這樣才會符合森林生態的常態。
就像前面提過的,很多耐陰性樹種發芽的時候,必須要受到不耐陰樹種的庇護,所以有不耐陰樹種在這邊也是有好處。但是過去的思維會認為看到不耐陰樹種會覺得這森林不好,這種想法可能要慢慢地把它改變掉,也就是說在森林的常態是一定會有不耐陰跟耐陰樹種並存在這裡,你讓這兩者都進來的時候,生物多樣性才可以獲得維持,而且整個循環才會比較完整。
過去臺灣很多的操作復育的人都有設定目標物種,這些目標物種可能都直接選擇耐陰性樹種,沒有提供不耐陰樹種的機會。但我覺得如果是要復育一個比較正常的森林生態的話,我們應該要在森林裡面提供不耐陰樹種生存的機會,也要提供它空間,才能夠維持一個比較大的生物空間。
提問:種子萌芽的關鍵:以前的植物生理學教科書中說,種子在黑暗中遇到水分就可以萌芽,不是這樣嗎?
不是這樣的。過去高中以下的教科書對種子需光這部分有點忽視掉了,教科書中說:種子在發芽這個階段有沒有光不是太重要,大部分是這樣寫,只有舉很少數的例子說,少數植物的種子是需要光才會發芽,大部分植物的種子發不發芽跟光一點關係都沒有。這種認知是錯誤的。
實際上在森林裡面,種子會不會發芽,有一大堆不耐陰樹種的種子發芽跟光度絕對是有關係。這類植物種子之所以會有這種偵測光度的能力,就是因為它希望種子一發芽出來就有很多陽光給它用,所以其實很多種類它的種子發芽是跟光有關係,只是過去的教材給了我們錯誤的訊息。
所以當這類種子休眠的時候,它就不會有任何作用,即使是下大雨有很多水分它也是不會吸水。對這類種子來說,光照就有點像是一把鑰匙,你如果沒有拿到那個正確的鑰匙它萌芽的機制就不會啟動。植物種子的生理其實很複雜,不同種類也有差異,沒有那麼簡單。
提問:種子休眠是因為沒有光,所以它就會進入休眠?
不是。有很多不耐陰樹種這一類的植物,種子傳播出來就會休眠,你不管怎樣它就自動進入休眠,好像它有一個定時器一樣,即使遇到合適的環境它也不長。有一票不耐陰樹種是這樣子,種子離開母體之後就自動進入休眠。
所以我們把這類種子取出來之後,要去打破它的休眠,要想辦法用很複雜的手續,這就是真的就是需要時間,有可能需要外在環境因子的某一種刺激,也會有其他很多的狀況。
種子生理其實遠比我們想像中要複雜,種子生理的書把耐陰樹種這一類的分成非常多的不同類型。
-第十章 結束-