本講義以文化大學森林暨自然保育學系蘇夢淮副教授於2023年為臺灣山林復育協會成員專業訓練課程內容為基礎進行整理編輯,作為山林復育種子教師及本會成員進修教材,以將學理知識對照應用,落實於山林復育行動。
課程重點:
1.進行森林復育時,植物補植方法的迷思:依靠現場灑種、自然萌發的方式雖然較符合自然的運作,但是很難成功。原因:
A.種子的萌發可能需要特定條件→種子的休眠性質。
B.種子可能會染病死亡或是被掠食。
C.小苗長成至小樹階段在森林更新過程經常是最具決定性。
i.小苗被掠食者吃食。
ii.因光度不足,當種子本身養分用完時,小苗會死亡。
iii.因根系太淺,遇長期乾旱會因缺水而死亡。
D.育苗後再補植可提高復育的成功率,但是種子的休眠性是育苗順利與否的關鍵。
2.種子的休眠性(Seed dormancy):
A.種子的休眠性可能是演化自避開種子剛離開母體時就馬上發芽可能面臨的風險。
B.外在休眠(Exogenous dormancy):造成休眠的因素位在胚(種子內的植物幼體)之外稱之。
i.物理性休眠(Physical dormancy):因種皮或果皮不透水造成。ex. 繖楊、山漆。
ii.結構性休眠(Mechanical dormancy):因種子被堅硬組織包覆造成。ex. 胡桃或核果類植物。
iii.化學性休眠(Chemical dormancy):胚之外含有會抑制發芽的化學成分。通常果肉為肉質多汁,或是種皮外覆黏性物質的植物會屬於此類,如柑橘、瓜類、番茄等。
C.內在休眠(Endogenous dormancy):因胚自身的因素造成休眠,稱之。
i.生理性休眠(Physiological dormancy):光因子,ex. 昆欄樹。溫度因子,ex. 萵苣 。臺灣的植物以此類居多。
ii.形態性休眠(Morphological dormancy):胚還未發育完全,種子便釋出(種子後熟現象) 。ex. 蘭嶼羅漢松、昆欄樹。
D.綜合因素:上述的休眠類別,是為方便研究所做的學理劃分,實際上同一種植物可能具有多類的休眠特性。
3.種子的儲藏性:
A.正儲型(Orthodox seed):種子可以保存在含水率0. 5%以下及0℃以下的條件。通常在長時間保存下還具有活性。
B.異儲型(Recalcitrant seed):種子通常在含水率30%以下的情況即開始失去活性,這類種子只能保存在低溫潮濕的條件下,且壽命通常不到一年。
C.正儲型種子通常比異儲型小,可在土壤種子庫中存活較久。
4.打破種子休眠的方法:
A.低溫層積:以低溫(通常為4℃)溼藏方式處理1個月以上(視種類)。此方法針對生理性休眠,為模擬冬季的環境。
易混淆名詞:春化作用,指花芽的分化機制。
B.光照:以適當光照射種子。此方法針對生理性休眠,模擬日照的環境。
C.除去種子外部抑制物質:水洗。此方法針對化學性休眠,為模擬種子的外部物質被雨淋洗或是被消化道消化。
D.破殼(內果皮、種皮等):磨破種皮,尤其是磨破水隙口(water gap)處,讓水分能進入到胚。此方法針對物理或結構性休眠,增快水分進入種子的時間。
E.文獻查詢方法:
i.學術方式:ex. Trochodendron(昆欄樹)+seed germination;同屬植物Styrax(安息香屬,臺灣野茉莉)+seed germination。
ii.非學術方式:ex.青剛櫟 發芽;ヤマコウバシ(白葉釣樟)発芽率。
5.遺傳多樣性:
A.遺傳多樣性是生物多樣性的一環,雖然重要但過去不容易探討。
B.遺傳多樣性來自於有性生殖與遺傳變異。
C.不同族群出現遺傳變異,稱為遺傳分化。
D.遺傳多樣性在生態上的重要性:
i.確保生物演化的重要基礎。
ii.增加物種存活的機會。
6.在地苗木:
A.復育時只考慮物種,雖符合物種層級的多樣性考量,但不符合遺傳多樣性的原則。
B.使用外地苗木,等於是引入外來的遺傳基因,會破壞自然演化的過程。附帶也可能會引入外來疾病(如白輪盾介殼蟲)。
C. 討論:復育上是否一定不能引入外來基因?
D.在地苗木的確認,由採種母樹開始。確認採種母樹的原生性,請參考本課程第2主題有關”剔除外來種與非當地的臺灣原生種”的部分。
「種子特性與遺傳多樣性」這堂課的主題設定在「種子特性」和「遺傳多樣性」,這是兩個部分,但之間有相關性,都關連到復育的基本部分,特別是「遺傳多樣性」在過去滿忽略的 。現在我們講的復育是「生態復育」,所以要去考量「遺傳多樣性」這一項。
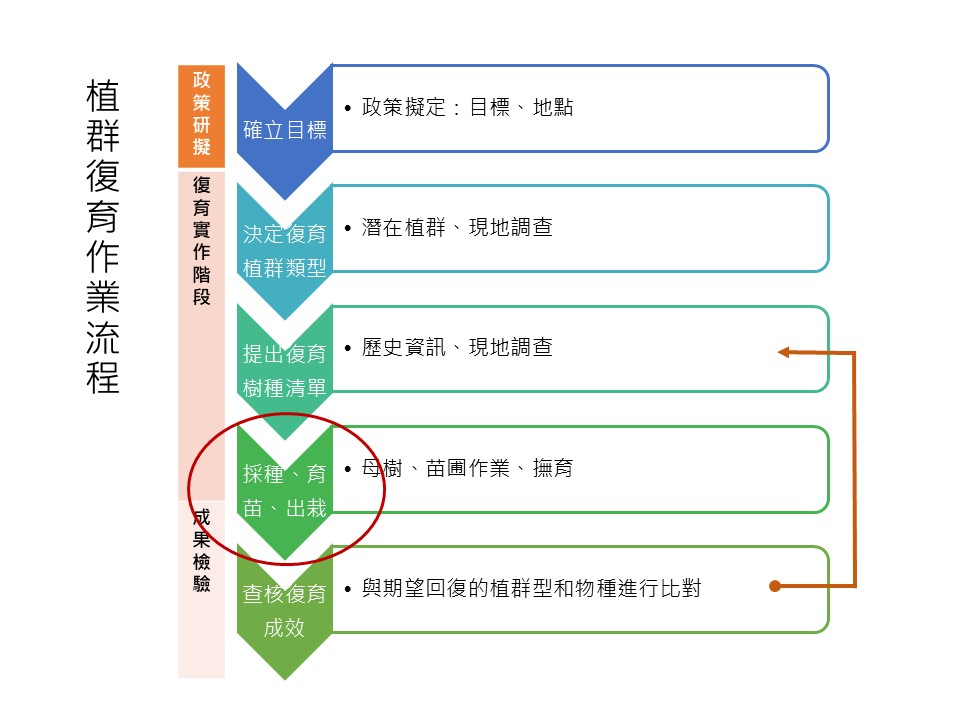
這一堂的主題「種子特徵與遺傳多樣性」,在整個植群復育作業流程裡面算是比較後端的部分。當我們把「復育樹種」決定之後,接著要「採種」、要「育苗」,育苗之後要「出栽」,所以是在很實作的這個階段,但是在這邊就會需要一些基礎的概念,也就是今天設定的「種子的特性跟遺傳多樣性」這個主題。
這堂課的主題有六個:一、 採用自然演替法進行復育的困境,二、種子的休眠性,三、種子的儲藏性,四、打破種子休眠的方法,五、遺傳多樣性,以及六、在地苗木的重要性。
第一個主題「採用自然演替法進行復育的困境」。所謂「自然演替法」,比如有一個生態破壞區,我們利用「大自然種樹」這種概念,我們都不要去做人為介入,這叫「自然演替法」。那麼,用這樣的方法進行復育的困境是什麼?
第二個主題是「種子的休眠性(Seed domancy)」,介紹種子在育苗過程裡面很重要的特性是種子的「休眠性」,我們可以嘗試從休眠性去了解一些種子,這個部分也會跟後面的主題會有關係。

第三個主題是「種子的儲藏性」。種子的「儲藏性」和「休眠性」不完全相等,但彼此會有關係,也跟我們在做復育工作在處理這些種子,以及選擇這些樹種的時候會有一些意義 。也就是說,你去了解種子的儲藏性質的話,有助於我們判斷我們在恢復的森林它植物的演替階段。
第四個主題是「打破種子休眠的方法」。 既然我們要育苗,就需要打破種子的休眠性。第五個主題是「遺傳多樣性」。最後,第六個主題是「在地苗木的重要性」。
1.進行森林復育時,植物補植方法的迷思:依靠現場灑種、自然萌發的方式雖然較符合自然的運作,但是很難成功。原因:
首先來看自然演替法的困境。我們在進行森林復育的時候,植物補植方法當然以前會用一些所謂的自然演替。後來就想,我們可以去現場灑種,加速它的生長,比如一些不容易傳播過來的樹種,我們直接現場灑種。但不管是自然演替或者是現場灑種,其實都很難成功,我所謂的很難成功,應該是說很難快速地達到我們預期的目標。
但是我們之所以要用人為的方式去介入森林復育,最重要的概念是希望加速森林演替的速度,加速恢復的速度,這是我們一開始有講的。如果我們不在意這個,我們願意等一兩千年,其實也沒關係,就放任它自然更新就好了。既然要加速的話,方法要對才可以正確地去加速。
這部份我們要來說明:為什麼現場灑種或者放任自然演替很難成功。
我用「森林更新過程」這張圖來說明整個森林的動態。「森林更新過程」這個階段是「雙向」的,雙向的意思是:從「成樹」或「小苗庫」做為開始都可以。我們從比較單純的,從「老樹」、「成樹」開始 。
在森林裡面,成樹就是大樹,它的未來會面臨兩個狀況:一個狀況是它會死亡,死亡之後當然它的空缺就會出現,可能別的植物會補進來。除此之外,成樹未來的另一個狀況是:它如果繼續活著, 它具有生殖能力,它就會繁殖,產生種子,種子落下來之後,在森林裡它可能會形成群生的小苗。我們通常在森林裡常看到:某一棵母樹下方有一群它的小苗,這個情形我們稱為「小苗庫」。
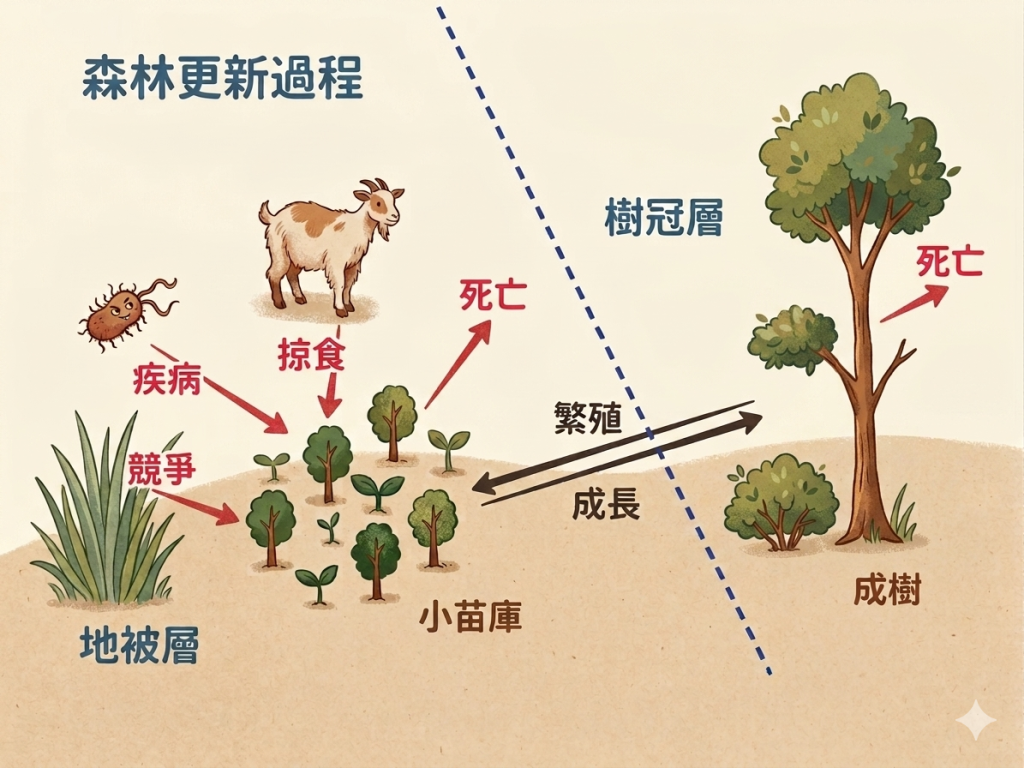
以常理來看,這一棵成樹它繁殖了很多小苗,但是你會發現這些小苗最後都不見了,這是合理的事,為什麼?因為一棵樹的壽命非常久,比如檜木會超過千年,如果每年只要它出現100棵小苗,這些小苗都存活的話,那一棵檜木它的繁殖力會有多強?它可以活1,000年的話,那是不是很恐怖?可是不可能有這麼多的樹活下來,所以它一定在某個狀況下,或是基於某些原因,它所繁殖的這些小苗大部分都死掉了,而且應該說絕大部分都死掉了。
事實上,就整個森林的穩定性來講,這棵成樹它只要在它的一生中有一棵小苗成功存活下來,然後長大,它就算沒有輸了。因為一對一,一棵成樹死掉還有一棵小樹出來補充,它的族群就不會減少了,這就是「穩定的森林」。
所以你想想看,它可能一年產生很多小苗,它產生了幾百年,但最終只有一棵活下來,其他都不見了。為什麼會不見?這是很多研究在探討的。
我們看到這張「森林更新過程」的示意圖,圖左下方這些小苗長起來之後,它可能基於某些原因直接就死亡,比如營養分不夠,養分用完了,它還沒有獲得光合作用的能力,它就死掉了;也有可能是被動物吃掉的;也有可能是得病,比如有一些病毒或是真菌這類的感染它,它就死掉了;它也會面臨其他的物種的競爭,因為在森林裡面植物之間互相競爭,競爭養分、競爭水分、競爭光度等等,所以樹的這些小苗面臨很嚴苛的篩選。
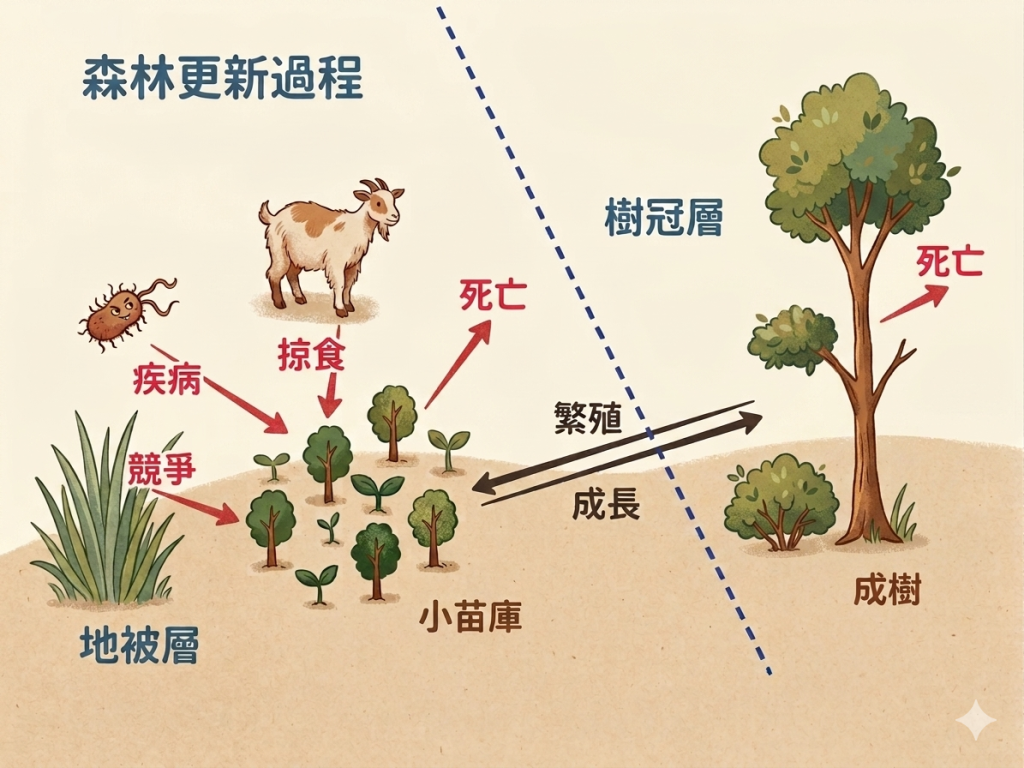
這邊舉例幾個主要狀況:
A. 種子的萌發可能需要特定條件 → 種子的休眠性質。
第一個原因是,種子的萌發可能需要特定的條件,也就是我們講的A.「種子的休眠性質」,沒有出現這個條件的話種子它不會萌發,所以它不會出現這些小苗。在第一關的時候,種子要出現小苗的時候就被篩掉了,連小苗都沒出現。
B. 種子可能會染病死亡或是被掠食。
第二個是,如果沒有出現小苗,除了休眠之外,還有一個情況就是B.「種子被吃掉了或是死亡」了。
C. 小苗長成至小樹階段在森林更新過程經常是最具決定性。
- iii. 因根系太淺,遇長期乾旱會因缺水而死亡
- i. 小苗被掠食者吃食
- ii. 因光度不足,當種子本身養分用完時,小苗會死亡
第三個是C.「小苗成長至小樹」的這個階段在森林的更新裡面也很決定性,就是前面提到的,因為小苗可能會面臨被吃,還有一些養分的供應問題,第三個是可能根系還不是很深的時候, 如果剛好遇到一個長期乾旱,地表都乾掉它就會缺水死掉,這樣的情況都可能會發生。
在不同的森林裡面,可能這些情況會是不一樣的。所以我在這邊舉一些例子跟大家說明。
這是我們臺灣自己的資料,在福山動態樣區做了大概8年的觀測,它的位置在北部福山植物園附近,一個25公頃的大的動態樣區。這個研究去去觀測冠層樹小苗的存活狀況。
在「福山動態樣區2003 ~2010年的冠層樹小苗存活曲線」這張圖中,橫軸是「時間」(年),從2003開始算,縱軸是「存活率」,是標記的小苗存活下來的比率。
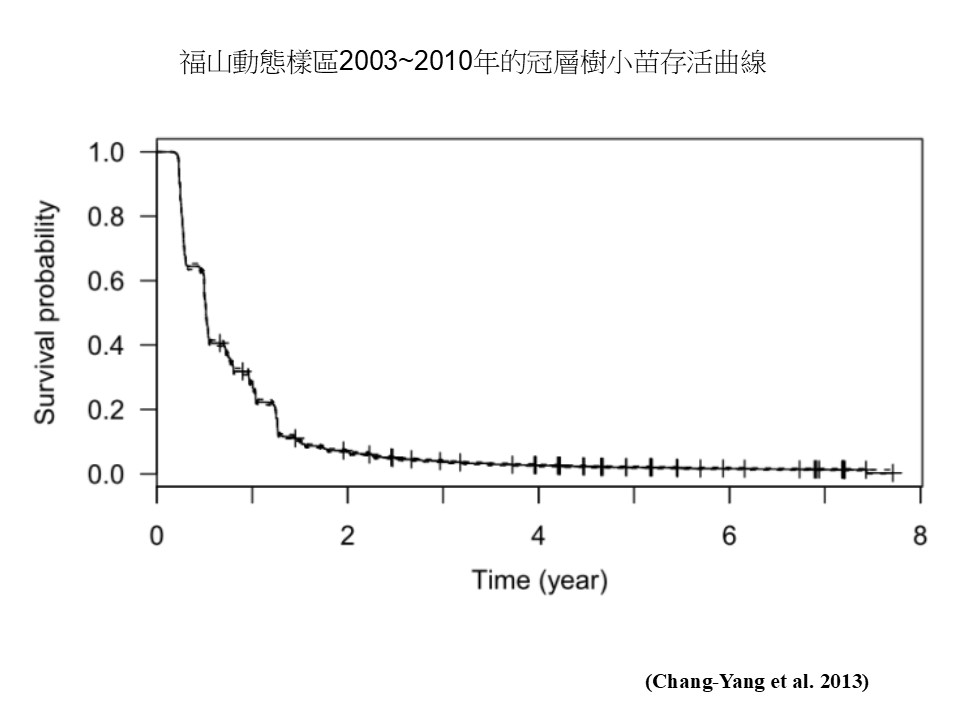
這個研究一開始監測的時候,以當時(2003年)為起點,所以每一棵小苗都是活的,存活率是100%。在第1年的時候,存活率已經下降到約20%,也就是第1年就有將近80%的小苗死亡。繼續監測下去,到第2年結束的時候,小苗剩下不到10%,但是之後(降幅)就趨緩了,可是一直到第8年,剩下的小苗幾乎全軍覆沒,存活率這一條線已經很接近0了。也就是說他監測的這些小苗裡面,在這8年來幾乎都已經全軍覆沒,只剩下一點點活下來而已,這就是一個森林的常態。它沒有跟我們講原因,只是就整個趨勢來看,小苗絕大部分都是沒辦法存活下來的,這跟我們在「森林更新過程」那一段提到的狀況是一樣的。
接下來這是我們研究室在玉山國家公園楠溪動態樣區所進行的小苗監測,做了大概有2年的時間。我們去紀錄每個樹種的小苗的狀況。
這張「楠溪動態樣區喬木小苗動態統計表」,第一欄是「物種名稱」,長葉木薑子、賊仔樹、假長葉楠等,第二欄是「曾掛牌的株數」,第三欄是「迄今存活數」,第四欄是「個體死亡的原因統計」,有「整株消失」、「乾枯」、「遭植食死亡」、「被壓覆或折斷」。最後是「死亡株數(死亡率)」、「曾受植食株數」。
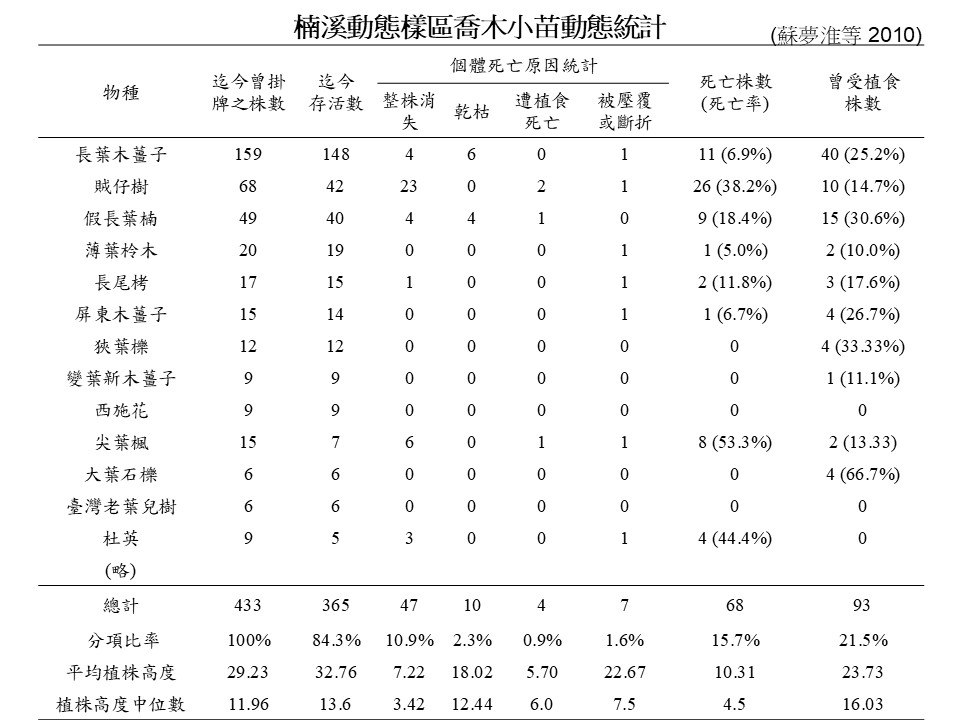
請看這張表倒數第四列,「總計」這個欄位,在我們紀錄的443棵裡面,在2年內死掉的,有47棵是整棵消失不見的,我們不知道原因。我們是每個月去一次,這個月它還在那邊,下個月就全都不見了,不知道是怎麼回事;但有一些它的屍體(殘餘)還在那裡,比如有10棵是乾掉的(i.e. 長葉木薑子6棵,假長葉楠4棵);有4棵是被吃死掉(i.e. 賊仔樹2、假長葉楠1、尖葉楓1);另外有7棵可能是被壓到死掉,總共是68棵死亡。也就是在 433棵當中,有將近 16% 的小苗在這2年的監測裡面死掉了。
回過頭來看,其實小苗整株消失的狀況裡面,我們判斷現場狀況是動物吃掉的,但是因為它很小,所以有些草食動物一吃就全部不見了,可能有些比較大的苗是上方(葉子)比較嫩被咬掉,死掉了,它下方的咬痕還在,所以我們才能判斷它是遭植食死亡的。這是在楠梓仙溪的情形。

我們當時用的方法是去做標記,每一棵小苗都用套環去做標記。所謂「整株死掉」是指植株不見、環也不見了;有一些個體比較小,甚至不能套環的,是插一個冰淇淋的木匙在旁邊直接寫編號,那一個板子也不見了。隔年我們加入自動相機監測的時候,發現有山羌、野豬、水鹿,那邊的草食獸很多,所以研判可能食物比較缺乏,草食獸數量又多,所以牠們就把小苗吃了,吃到連只剩木匙的那一段牠也吃掉,因為怎麼找都找不到木匙的殘留物。
這個例子是說,楠溪動態樣區這邊野生動物多,所以小苗的篩選的主要因素是被動物吃掉。而前一個例子福山動態樣區的研究,他是比較全面地( 大尺度的)去看小苗存活率,沒有去探討小苗死亡的因素。
我們看澳洲桉樹(桉屬eucalyptus,或稱尤加利屬)的例子。在澳洲這種比較乾旱的環境裡,供水量以及溫度是影養種子萌發時間點的主要因素。在西澳地區,許多的桉樹種類因為氣候(特別是年雨量)以及土壤類別,呈現出明顯分布模式。
澳洲的這個研究選取了4種桉樹種子做實驗,在「4種澳洲桉樹種子萌發率對於土壤水的反應」這張直條圖中,4種圖樣分別代表不同種類的桉樹,縱軸是種子發芽率(%),橫軸是土壤水(水勢),單位是壓力MPa (百萬帕),負值越大表示土壤水越少。
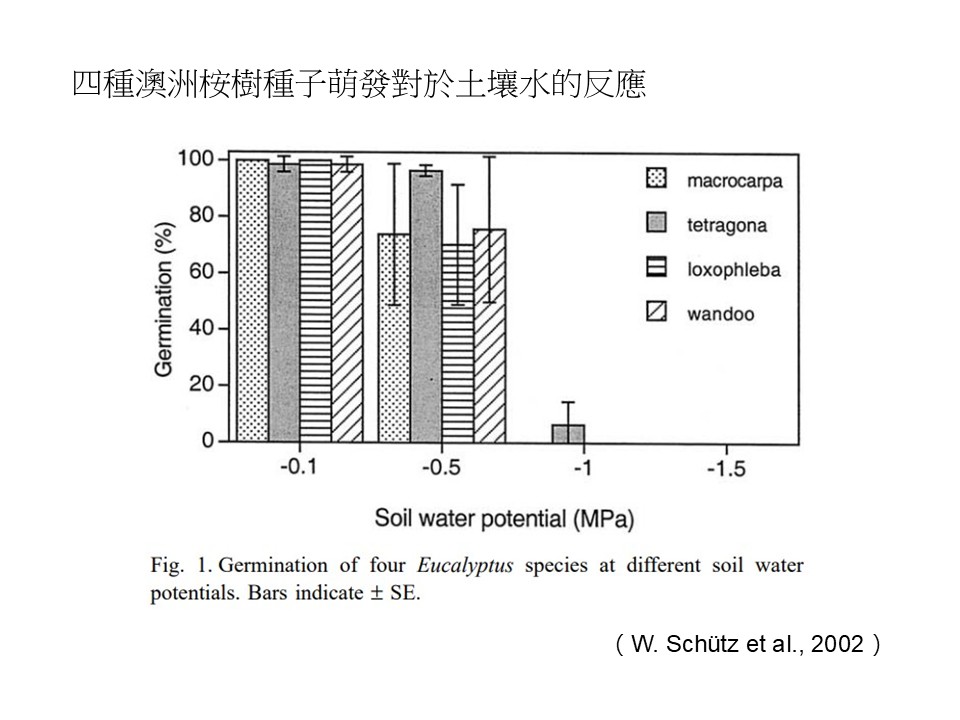
研究結果顯示,幾乎所有的種子在-0.1MPa時都會發芽,而水勢在 -0.5MPa的條件時,發芽率稍微降低。土壤水勢為 -1 MPa時,僅剩Eucalyptus tetragona這種的種子能夠發芽,但是發芽率不到10%。當土壤水勢低到-1.5 MPa時,全部4種都無法發芽。
在臺灣的話,目前動態樣區都設在雨林,水分很充足的環境,所以目前看不出來水分對我們的小苗是一個很大的限制,因為水分供應在我們的野外算是很穩定。
D. 育苗後再補植可提高復育的成功率,但是種子的休眠性是育苗順利與否的關鍵。
2.種子的休眠性(Seed dormancy):
A. 種子的休眠性可能是演化自避開種子剛離開母體時就馬上發芽可能面臨的風險。
在種子的休眠性這個主題一開始,先講「種子休眠的分類」。種子「休眠」的定義是,種子傳播出來離開母樹之後,它可能會經過一段時間才會啟動它的萌發機制,還沒啟動之前的那段時間我們稱為「休眠期」。休眠期可能很短也可能很長,狀況非常多。一般認為種子離開母樹之後會產生休眠,是一種保護機制,它會先睡覺,過程中等待,等待環境好的時候,它發現有訊號在-環境變好-它就萌發。它一離開母體如果遇到壞的環境卻還是發芽,它可能就完蛋了,所以種子休眠這是一種演化上的一個機制,一般都是這樣去解讀。但要要獲得這樣的特性,它的原因就變得很複雜,不同的科,或者是不同的環境的物種,休眠的方式可能就會有些不一樣。
我們以這張圖為例,這個種子它是「豆科」的,例如「花生」,或是「豌豆」、「黃豆」這類的,你可以把種子剝開變成兩瓣,裡面你會看到小小的這一個是「胚」,就是植物的幼體。
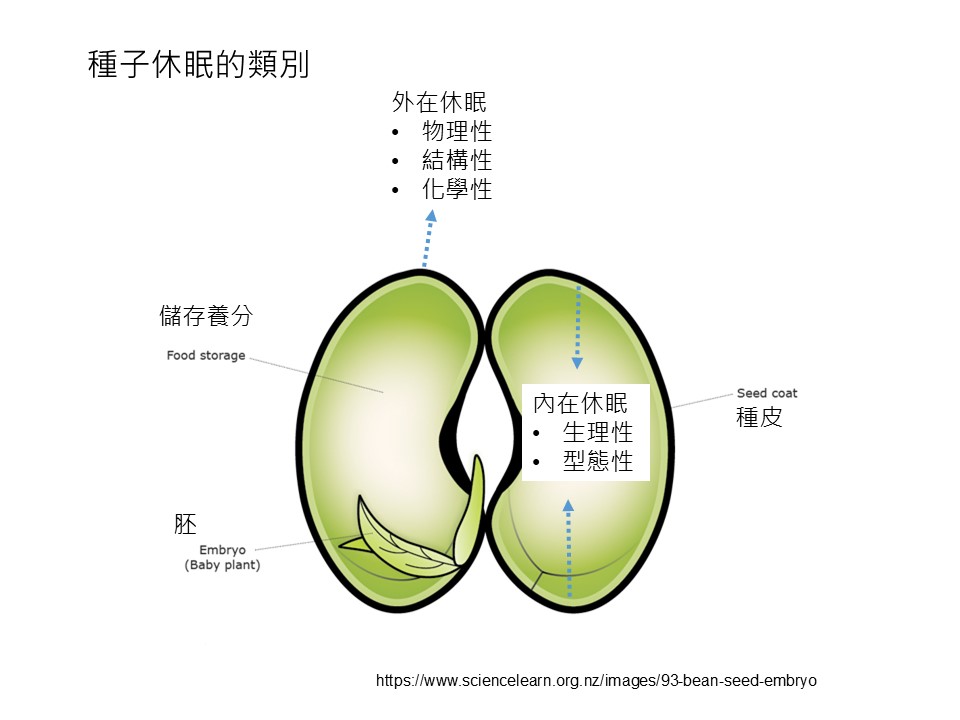
我們講的「發芽」,它的定義是「胚根」伸出「種皮」之外。如果胚它一直躲在種子裡面,還沒有伸出整個種皮之外,都定義它還沒有發芽。
「胚」它會連接的主要的部份是種子「儲藏養分」的部分,所以你可以預期,越大顆的種子儲藏的養分愈多,並不是胚愈大。這是大型種子和小型種子主要的差異。
「胚」以外的種子的部分幾乎都是以「儲藏養分」為主。
當然它還有另外一個功能,在某些植物裡,這些儲藏養分的是「子葉」,所以在萌發之後「子葉」會開始行光合作用,也幫它供給養分,除了本身儲藏的養分之外,它也可以自營生活供給養分,這是儲藏養分的這些構造。
再來是「種皮」,就是包覆種子的最外層。
以上,我們把種子分成這三個部分-胚、養分儲存、種皮。
B. 外在休眠(Exogenous dormancy):造成休眠的因素位在胚(種子內的植物幼體)之外稱之。
i. 物理性休眠(Physical dormancy):因種皮或果皮不透水造成。ex. 繖楊、山漆。
我們定義「種子休眠」有兩大類因素,一類是「外在休眠」,一類是「內在休眠」。兩者差異是「外在休眠是在種皮之外發生」,「種皮以內的稱為內在休眠」,這是非常人為的切分法,因為這個區分的關鍵點是在「種皮」,種皮的狀況沒有那麼一致,所以可能有一些內在休眠是牽扯到種皮的,但是有一些外在休眠也是牽扯到種皮的,「種皮」這層變成一個模糊空間。
「外在休眠」分成三種:「物理性」、「結構性」、「化學性」。
「內在休眠」分成兩種:「生理性」、「形態性」。
在某些休眠的分類裡面,它不是這樣分,因為各種分類法的著眼點不一樣,分出來的名詞就會不太一樣。但是我們今天先用這個版本。
「外在休眠」裡有一種是「物理性休眠」,是因為「種皮」、「果皮」、「果肉」的部份具有不透水層構造,水分無法進到種子,就形成「休眠」。
照片中這是「山漆」,果皮表面很油亮,果肉裡有很多蠟質,也就是油。油和水是不互溶的,當果肉或果皮有很多油脂的時候,水是無法進去種子,造成的問題是什麼?種子無法吸到水,它就不會萌發,因為它需要吸水開始產生它的生理反應,才會細胞分裂長大,所以種子吸水這件事對種子萌發很重要。

提問:既然「種子吸水」那麼重要,它為什麼要讓吸水變得這麼困難?
答:這就是剛提到的,在長期的演化裡面,當種子它傳播出來的時候,如果是長期面臨一個不好的環境,對它(的存活)來講是不利的。所以如果它的種子很容易萌發,小苗一下子就長出來,它可能會馬上遇到不好的環境,它就會死掉。
我們可以想想看,在長期演化裡面,這些物種為什麼它的整個脈動的頻率會很接近它的環境?因為不接近它的環境運作的這些物種都死掉了。
一個物種為什麼會接近這樣的脈動,有些不是?因為有遺傳多樣性,就是我們第二個主題,所以它有一些差異。在這些差異下這個環境去篩選它們,不適合環境的就被篩選掉了,適合的它就留下來。
你看到這個「山漆」,為什麼它的果實變成這樣有很厚的油脂?,可能說明種子剛離開母樹的時候,它遇到的環境是不好的。
「山漆」是臺灣的物種,果實成熟期是秋冬,在9月、10月差不多成熟,要進入冬天了,這就是解答。所以它如果這麼傻在冬天萌發,它會面臨冬天的惡劣環境,也就是「低溫」。也許我們不會覺得冬天溫度有多低,可是山漆在臺灣它是分布到海拔2,000多公尺的物種,冬天可能是個位數的溫度,甚至有時會0度以下。其實「山漆」是在整個東亞北邊都存在的,臺灣是這類物種分布的最南邊。所以想想看,再往更北的地方,冬天是會下雪的。我們臺灣很多的物種是冰期的時候過來的,其實是代表北方溫寒帶的這些物種,它的老祖宗是遇到會下雪的環境,到臺灣這邊比較溫暖,可是它這個特性還保留著,因為演化所獲得的行為能力,它不是隨便改變一下環境它就會跟著改變,沒有那麼快,可能它的尺度是好幾百萬年才會改變這個習性。所以臺灣的山漆跟它的北方這些祖先分開的時間可能是幾十萬年的尺度而已,所以它還保留著北方祖先的特性,它要度過這個冬天所以演化出不透水層的構造。
這個不透水層構造終有一天會打破。怎麼打破?外在環境有很多病菌、微生物,會慢慢地把它的油脂吃掉、分解掉。微生物會去找有機物去分解,可能剛好在果實表面上,它就慢慢去分解,分解之後形成孔洞,如果剛好有水來就滲進去了。這個油脂不會一直存在,會慢慢被微生物分解掉,所以它需要時間。也許這分解的時間剛好是春天來的時候,春天是雨季,它就被配合這段時間就可以順利萌發。
這種休眠的狀況,是因為水分沒辦法很快的進入種子,進入到它的胚,隔著不透水層,稱為「物理性休眠」。
提問:「無患子」是不是也是同樣的機制?無患子的綠色落果,剝掉果皮播種,在當月就會發芽,但如果是褐色果實就要隔年春天才會發芽,所以是不是果皮綠色時期種子還沒進入休眠?
答:「無患子」的概念類似「龍眼」。我們吃的龍眼果肉其實不是果肉,是「假種皮」,那是種子外的一層構造,吃完果肉裡面剩下一顆黑褐色的種子,黑褐色的部分就是種皮。
龍眼的種皮很油亮,完全成熟的無患子的種皮也是很油亮。我在想,在果皮綠色還沒有完全成熟的時候就把它剝掉,它種子的種皮可能還沒發育完全,可能本身就有很多孔洞,水很容易進去。我在想可能是這樣,但我沒有特別觀察過。
你可以比較,綠色果實的種子是不是比較柔弱。如果是的話,代表它還沒有發育完全。如果是發育完全的種子的話,種皮應該滿硬的、很厚實,水不容易進去,就需要去打破不透水層。
綠色果皮就播種的話有一個風險,我們採的是未成熟果,它的胚還沒有發育成熟的時候,它可能是比較柔弱,也就是培育出來的小苗雖然你看到它萌發但它的競爭力是很弱的,概念上有點是早產兒。比較正規的方式應該是等種子全熟的時候,再用一些方法去處理它,可能會比較好。不過有些物種很厲害,比如,我認為「龍眼」可能就比較不怕這件事。
以前我們在算都會區入侵植物的入侵性,有一些臺灣的百大入侵物種的物種,大家都有看過。可是我們算起來,龍眼是臺灣入侵植物裡面,入侵指數最強的,它在我們低海拔確實很嚴重。我們低海拔森林很多龍眼樹,而且這些龍眼樹可以自己活下來,但因為它是經濟樹種我們不太覺得有什麼問題。不過你如果從生態面去看,龍眼是滿令人擔憂的,甚至比陰香還厲害。龍眼果實落到地上,種子就可以長了,它不需要經過動物的消化道。消化道是一個加速的過程,不過絕大多數的物種不需要經過消化道。
我們看另外一個物種「繖楊」,又叫「恆春黃槿」,它是長得像黃槿的一種植物,也是海岸的「錦葵科」植物。「繖楊」在臺灣過去被認為是比較稀有的植物,因為它的原生地有一些生態破壞。後來「繖楊」有在做一些人工栽培。
照片中是「繖楊」的果實,這是「蒴果」,裂開之後裡面是它的種子,它的不透水層主要是在種子上,種皮有不透水層,種皮有一個比較薄弱的點,稱為「種皮合點栓」,這個點比較容易被水穿透,當種子泡水一段時間,種皮合點栓爛掉了,水就容易進去。就像管道裡面有一個壁比較薄,水不斷地摩擦,壁破掉了,水就進去。這也是一種「物理性」的休眠。
「山漆」的不透水性是在「果皮」,「繖楊」的不透水性是在「種皮」。
提問:所有種子都有「種皮合點栓」嗎?
答:最重要的不是「合點栓」,這個部分主要講的名詞是「水隙口」,水會通透到種子裡的(構造)叫「水隙口」,這個口的位置在「種皮合點栓」。
提問:黃槿發芽率不是很高,國外文獻有講黃槿種子的處理是泡鹽酸、或是用砂紙磨、用熱水燙,是否就是要處理掉它的厚壁細胞?
答:對,應該也是類似這樣。你們有試過泡水泡很久的方式嗎?我認為這類的具不透水層特性的,泡水泡很久應該有效。
「山漆」要進行休眠是因為「溫度」。「繖楊」它是恆春海岸的物種,它的來源是熱帶海洋,它為什麼也有休眠機制?因為它要「海漂傳播」,所以它必須要很輕、要有一個不透水層在它種皮上。為什麼「繖楊」的不透水層不是在果皮上面,而是在種皮上?因為「黃槿」是「蒴果」,「蒴果」成熟時會裂開,把種子釋放出來。所以如果它的不透水層是在果皮上的話 ,果實成熟時開裂,海水一進來,種子可能就完蛋了。所以它的不透水層是在種皮上面,用種子去海漂,它的不透水層表層要讓水分沒辦法通透,所以表層的油的成分濃度會很高。
為什麼它一旦上岸就可以解除?重點就在「淡水」,因為淡水接觸到種皮。雖然它是油脂水沒辦法穿透,可是淡水裡有很多微生物會把油分解掉,這時候它就開始有通透性,淡水就會進去。但是海水不會有這個問題,因為海水裡沒有分解油脂的微生物,海水鹽度濃度很高,所以一些小型的微生物是沒有辦法存活的,所以在海水裡種子有點像是在保存的狀態,等它一上岸的時候,淡水的微生物就把種皮分解掉,淡水就順勢進去,它的胚就開始萌發。
我覺得下次可以測試泡淡水。泡水不要泡滿,因為種子都在水面下可能會沒辦法呼吸,水裡氧氣不足,泡太久種子會缺氧而死。所以要有點半泡著,上層還是要讓它接觸空氣,下面有水慢慢浸溼、把它浸透。
蔡智豪:我們這方法是沙藏,用沙子的溼度去埋種子。
蘇夢淮:這也是一種方法。沙藏跟泡淡水不一樣的地方是,沙藏沒有光。我們等下會講到一種休眠是跟光有關的。如果這個種子它對光這個條件不在意的話,會成功。但如果這種子是需光的種子,要用光去解除它的休眠,你用沙藏法會失敗,因為沙藏是蓋在沙下面,種子沒有光照,但是泡水是不會擋住光線的,算是比較萬用的方法。
黃香容:所以我們可以模仿它被海帶到海岸邊的時候。
蘇夢淮:講到重點了,我們後面有講,所謂打破休眠就是要先了解休眠的機制,我們了解之後就是模擬自然界打破它的方式去打破。
ii. 結構性休眠(Mechanical dormancy):因種子被堅硬組織包覆造成。ex. 胡桃或核果類植物。
外在休眠的第二種叫「結構性休眠」。以「臺灣胡桃」為例,分布在臺灣中低海拔比較局部性地區,比較乾旱一點的地方,它有滿大的族群;臺灣比較大的族群在中部的山區,梨山;還有在東部的低海拔。

這張照片裡是臺灣胡桃的果實,外面果肉吃完之後會有一個硬核(右圖),這個硬核叫「內果皮」,不是種皮,種子在內果皮形成的這個空腔裡面,這類的果實叫做「核果」,例如「李子」、「桃子」也是這樣,裡面會有一個硬核,這類屬於核果,硬核是內果皮。
「臺灣胡桃」的種子為什麼不容易發芽?因為內果皮很硬,水是能夠進去因為有一些裂隙,把它泡水就可以。可是因為它的內果皮太厚太硬了,胚是軟組織,胚它長大之後它沒有辦法推開內果皮,所以需要不斷泡著水,慢慢軟化,而且從乾濕交替之間,內果皮有一些接合縫會裂開,胚就有機會伸出來,把整個果實內果皮撐開。所以臺灣胡桃休眠的主要原因是內果皮太厚太硬,這叫做「結構性休眠」。
「結構性休眠」感覺上跟「物理性休眠」有點像,差異是在於物理性休眠種子是不透水的,結構性休眠種子是因為很厚很硬的構造把種子包起來。
提問:「降真香」的休眠是屬於那一種休眠形式?
答:降真香比較是「結構性休眠」。降真香也是核果,協會要育苗最簡單的作法就是把它破殼。
提問:後來放一年六個月就大量萌發。
答:因為降真香這種就是需要長時間,去把內果皮弄軟,或是讓它有點在乾溼的過程裡面去裂開。
提問:把它輕輕敲裂一點,再把它放著,這樣可以嗎?
答:可能需要一些工具,大家知道有一種工具叫胡桃鉗。胡桃鉗的好處是夾下去內果皮會破掉,但裡面的胡桃是完整的。你可能要找到這種工具,可以省力地壓下去,殼破掉但是裡面的胚不會傷到。胚非常軟,它都是薄壁組織,只要不小心讓它有機械性傷害就會死掉,因為它那時候的胚很弱它會感染,只要有一些破口,外在的病菌就會進去它就死了。
所以做破殼,動作要很小心,要找到合適的工具,讓你很有效地破殼又不會傷到胚。我在網路上有看到一種方法是用老虎鉗,老虎鉗有個洞,在開口墊個合適高度的橡膠墊,夾下去殼會破但胚沒有受傷,大家可以參考看看。
iii. 化學性休眠(Chemical dormancy):胚之外含有會抑制發芽的化學成分。通常果肉為肉質多汁,或是種皮外覆黏性物質的植物會屬於此類,如柑橘、瓜類、番茄等。
再來是外在休眠的第三種「化學性休眠」,種子外層有一個需要消化的物質。
我們以「哈密瓜」為例,通常瓜類的果實切開之後有很多種子。這個種子有時候很討厭因為它很黏,黏在你的手甩都甩不掉,到處粘。種子外面這層黏性物質含有一些化學成分會抑制種子休眠。為什麼?這也是演化的結果,推測是因為這是動物在吃的,動物牠把果實整個吃掉,這些種子被吃到肚子裡,消化道裡的消化液就會把抑制物質除掉,當種子隨著糞便排出的時候,種子表面已經沒有化學物質,它就可以順利萌發。有人認為這種演化就是配合動物攝食的習性,因為這種種子經過消化道沒有被消化,之後跟著糞便排出來,這個種子是在糞便堆裡面,它有養分,反而對種子萌發來講是有利的。
提問:哈密瓜被吃的時候,它怎麼知道它的種子會經過這隻鳥的胃,然後演變出這個黏液?
答:它不會知道。我們可以想像一個情境,很古早的時候,瓜科的祖先它有很多的後代,有的後代有這個能力,有的沒有。具有這個性質的,它可能在後代的競爭裡面比較強,因為它存活率高。怎麼說存活率高?因為它種子掉出來之後要萌發的時候,除了有本身的養分,還有動物糞便的養分;另外一批它沒有這樣的機制,它只能靠自己,所以那批死亡率可能就會高,比較容易被環境篩選掉。所以種子具有黏液的瓜類應該就是整個演化篩選之後的結果。因為祖先已經具有這個特性,遺傳到後代的時候可能即使已經不同種,但這個性狀還被保留著,所以一直有這樣的特性。大概我們在演化都這樣子解釋。
我們必須知道,演化它其實是沒有目的性的,因為後代的遺傳基因的組合是隨機的,所以父母親沒辦法控制後代的基因型,唯一能控制的是各出一半基因給後代,但要出哪一半基因不知道,也沒辦法控制,就像我們講生男生女是沒辦法控制的。但是因為後代是遺傳基因不一樣,所以它有多樣性,有的表現這樣,有的表現那樣,最後環境演化把它篩選出來。
提問:是不是像「破布子」、「榕果」、「構樹」,也是屬於這一類的?「破布子」超黏的。
答:有這種黏性物質的通常會這樣。我舉例,在我們復育工作裡最常遇到的就是「榕屬」植物。榕屬植物的種子都會黏,有些特別黏,像「愛玉」那種,愛玉可以洗出果膠。果膠對種子來講雖然是不好的會抑制它發芽,但對其他生物來講是很營養的食物。這是我們講的「化學性休眠」的機制。
以上這些都是外在的休眠,因為它是表現在種皮以外的。
C. 內在休眠(Endogenous dormancy):因胚自身的因素造成休眠。稱之。
接下來我們看「內在休眠」。第一個是「生理性休眠」,我們這邊講的有幾種主要的,一個是光一個是溫度,這是最常被提出來的。
i. 生理性休眠(Physiological dormancy):光因子,ex. 昆欄樹。溫度因子,ex. 萵苣 。臺灣的植物以此類居多。
「 內在休眠」第一種是「生理性休眠」,「光」和「溫度」是最常被提出來的,是胚自己生理的狀況。
有些種子需要有光,有些種子它一定不能有光,需要在全暗的地方才會發芽。基本上需光的種子比較多。
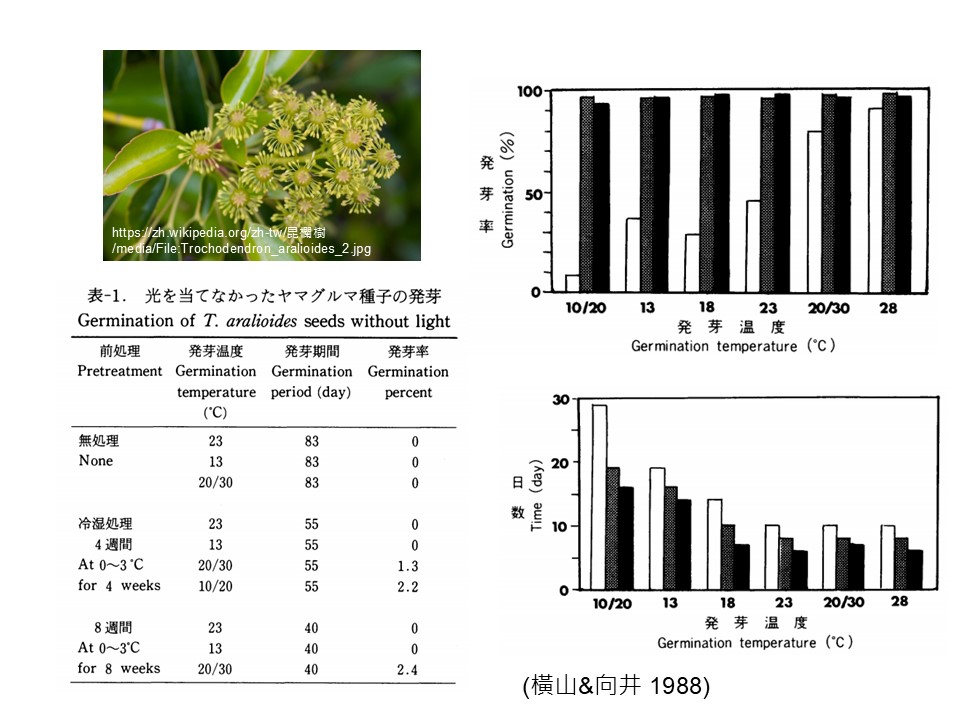
我們舉「昆欄樹」(Trochodendron aralioides)的例子。日本他們有做過一些實驗-昆欄樹的種子在無光條件下發芽情形,讓種子在黑暗中,他處理了很多不同溫度,因為他要測試昆欄樹種子的生理休眠特質,不照光,全部用「溫度變化」去控制, 結果所有的處理,種子幾乎都沒有發芽,只有少數1.3%、2.2%的狀態,很少。基本上你可以把它視為後代的變異,意思是後代有些改變了它本來的性質。
就像前一個提問到關於物種的演化。後代總是有一些變異,也許今天又來一顆小行星撞擊地球,地球又全部黑暗,這種很奇怪的這些(1.3%、2.2%的)後代它可能就會活下來,其他需要光的後代全部都死光。只要這些變異的後代活下來,它又可以慢慢地壯大。
這個研究顯示「昆欄樹」它發芽一定要光,在有光的狀態下,溫度對它沒有太大的不同。我們看右上圖,這是用變溫處理,10℃、20℃,或是從13℃一直到28℃,他都去做實驗,結果它的發芽率都差不多,都將近100%,所以最重要就是要照光,這是「昆欄樹」。
但是雖然發芽率差不多,但是發芽的時間不一樣。我們看右下圖,高溫會縮短昆欄樹發芽的時間。種子種下去,如果持續低溫,15天以上到將近一個月它才會發芽,可是如果在18℃甚至23℃,它可能10天左右就會發芽。所以溫度不會影響發芽率,但是會影響它發芽的速度。
這我們要怎麼解讀呢?因為「昆欄樹」是溫帶植物,溫帶的冬天是冷的,春天也是冷的,只有夏天才是溫暖的,所以這是在協助它在夏天發芽,太冷的時候它先不要發芽。比如在10℃的時候,是春天左右,讓它延後發芽時間;到夏天叫它要趕快發芽。就是種子遇到不同溫度,它反應不一樣。
這種情形反應的就是「生理休眠」,種子在黑暗的時候不會發芽:什麼時候不會發芽,因為比較北方的溫帶,冬天日照很短,這段時間它是不會發芽的。要等到某個程度,照光的時間足夠的時候它才發芽,這個時候溫度也拉高了。所以從這個數字來看,最好是夏天發芽,對它來講是最合適的生長季。
提問:白色長條是甚麼意思?
答:可能是控制組或對照組之類,要看原PAPER。
提問:這跟先驅植物有關係嗎?
答:有關係,如果是「光」這個條件的話,先驅樹種通常是光誘導的,它的「『休眠與否』會控制在『光』(+這個條件)」。例如「山黃麻」、「臺灣赤楊」。「臺灣赤楊」是崩塌地的物種,林務局在崩塌地種很多臺灣赤楊來做水土復育。「臺灣赤楊」就是典型光誘導的(+物種),光線充足的話它就會發芽,光線不夠它就不發芽,它這也是「生理性休眠」的展現。
D. 綜合因素:上述的休眠類別,是為方便研究所做的學理劃分,實際上同一種植物可能具有多類的休眠特性。
我跟大家補充,其實「休眠」的這些(+類別),是我們人為硬把它切出來的,實際上很多種子都是具有多重的休眠性質。例如「昆欄樹」它同時有「形態休眠」,也有「生理休眠」,昆欄樹的胚它在一開始沒有發育完全,也是需要經過一段時間(+繼續發育)。所以說,昆欄樹的種子你一開始就把它丟到有光的環境,溫度也合適,它也不會那麼快就長,因為它的胚還沒有完全發育好,所以,(+植物種子的休眠特性)通常是綜合性的。
3.種子的儲藏性:
A. 正儲型(Orthodox seed):種子可以保存在含水率0. 5%以下及0℃以下的條件。通常在長時間保存下還具有活性
B. 異儲型(Recalcitrant seed):種子通常在含水率30%以下的情況即開始失去活性,這類種子只能保存在低溫潮濕的條件下,且壽命通常不到一年
C. 正儲型種子通常比異儲型小,可在土壤種子庫中存活較久
4.打破種子休眠的方法:
A. 低溫層積:以低溫(通常為4度C)溼藏方式處理1個月以上(視種類)。此方法針對生理性休眠,為模擬冬季的環境。
易混淆名詞:春化作用,指花芽的分化機制
B. 光照:以適當光照射種子。此方法針對生理性休眠,模擬日照的環境。
C. 除去種子外部抑制物質:水洗。此方法針對化學性休眠,為模擬種子的外部物質被雨淋洗或是被消化道消化。
D. 破殼(內果皮、種皮等):磨破種皮,尤其是磨破水隙口(water gap)處,讓水分能進入到胚。此方法針對物理或結構性休眠,增快水分進入種子的時間。
E. 文獻查詢方法:
i.學術方式:ex. Trochodendron(昆欄樹) +seed germination;同屬植物StStyrax(安息香屬,臺灣野茉莉)+seed germination。
ii.非學術方式:ex. 青剛櫟 發芽;ヤマコウバシ (白葉釣樟) 發芽率。
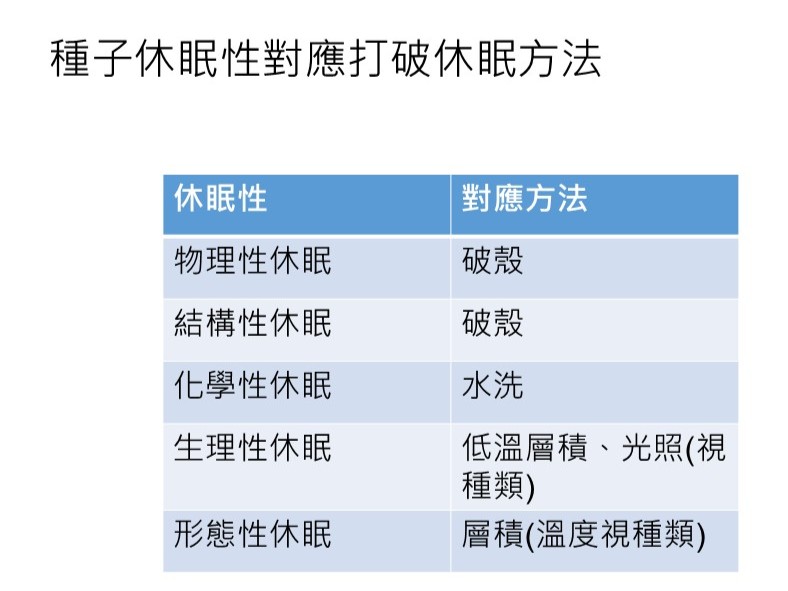
我們從「種子休眠性對應打破休眠方法」這個表格,整理出打破種子的休眠的對應方法。如果我們育苗都用天然的方式(去做)你可能會很挫折,要到那個(物種的發芽)條件要怎麼去模擬?
「物理性」和「結構性」的話,基本上就是「破殼」,但是你要知道要處理的是「果實」的部分還是「種子的種皮」的部分,要去針對它來打破。「破殼」基本上只要讓它裂開或是敲掉一角就可以了,不需要把整個殼都剝開取出種仁,種仁拿出來其實很容易傷害到它,只要有破就可以了。
提問:有裂縫就好嗎?
答:裂縫要看裂多大,裂開一角比較好,這樣胚比較容易出來。只列開縫隙的話,因為胚根很軟,結構休眠或物理休眠的,它的核是比較硬的殼,如果只是一個裂縫,有時胚根要出來的過程它會失敗。如果裂縫是一個面的話,它就比較容易鑽出來。
「化學性休眠」基本上因為它是種皮外有化學物質抑制,所以就用水洗的方式,也就是泡水。比如泡一天就要換水,不然抑制物質會累積在水裡,會造成抑制,所以要不斷地換水。
「生理性休眠」就要看它的狀況,所謂的「低溫層積」,就是放冷藏4℃,或5℃,(+同時)要保濕。通常有生理性休眠的物種,它通常是沒辦法耐乾旱。我們之後在種子儲藏的特性裡面會講到,種子的含水的比率有一些關係。「生理性休眠」也可能是用「光照」(視種類)來促進它(發芽)。
「形態性休眠」就是用「層積」,溫度可能要看種類。熱帶物種可能不要用低溫,「常溫的層積」只是保水儲藏;但是「低溫層積」就要進冷藏。所以「形態性休眠」要看這些物種的特性來決定。
提問:「羅摩科」用哪一種休眠?
答:我沒有看過休眠的例子有講到羅摩科植物。(形態性休眠會不會跟吃它的動物有關?)
這些休眠類型都被歸為一個因素,就是要度過不好的環境,只是它方式不一樣,所以才歸為不同的休眠方式。 比如它的表現,剛好是形態性休眠這種方式這樣。
提問:因為有些蝴蝶的幼蟲,牠門吃食草的時候會整株吃光光,幼蟲一出來的時候是超多的,會把植株整個都吃掉。華他卡藤那種的也是,如果它的幼苗下去,長出來的時候,那些昆蟲也還沒有離開,它那時出來也會被吃光。所以我在猜測會不會是這樣子(有形態休眠)。剛才提到的蘭嶼羅漢松也是,有一種蛾也是很愛吃,吃到整株都沒有。能活下來就是必須要蝴蝶化蛹,整個世代都離開之後,它再出來才不會死。
答:這樣你就要看吃它的這些昆蟲牠的季節的脈動,跟種子的...。蘭嶼羅漢松這是比較特別,我是沒有看過昆蟲會把牠的食草整個吃死的。(蔡:應該是不會被吃死,只是吃光光而已)
蘭嶼羅漢松是會死,但是它的死很特別,它第一次被吃的時候不會死,葉子都被吃光了,吃完之後它會再長新葉,同時長新葉,假設它在長新葉那階段被吃第二次,因為它的養分用完了,它就會死了。接續被吃它就會死掉,如果沒有的話,它就又可以活得好好的。
我們在野外看,之前在楠溪觀察,看到有一種赤楊金花蟲,它大發生的時候也會把臺灣赤楊的葉子吃光光,整片赤楊林都沒葉子。赤楊是落葉樹,但是我們看到它是在夏天沒葉子,就是被蟲吃光了,吃完之後赤楊又會長新葉,可是金花蟲不會再一次大發生。牠再第二次大發生可能是很長很長一段時間以後。所以橙帶藍尺蛾它是比較特別,牠竟然會在吃完蘭嶼羅漢松之後又來一次大發生,就是它已經把食草吃乾淨了,不久又來一次大發生,怎麼會有這種物種?這很奇怪,這可能要看做昆蟲的人他們有什麼的解釋。一般來講不會吃死。橙帶藍尺蛾是我唯一看到一種會把它的食草整個吃到光光的這樣。
蔡:牠是自我控制牠的族群數量,因為牠自己也自殺了。
蘇:對,可是牠並沒有滅絕。有趣的地方是牠並沒有滅絕。等於是牠的周期很短,我是比較沒辦法想像。不過這是另外的問題。
5.遺傳多樣性:
我們提到,「遺傳多樣性」是「生物多樣性」的其中一環,我們講「生物多樣性」都會講「生態系」,生態系就是有針葉林、有闊葉林或是有沙漠、草原,這些叫「生態系」。
在同樣的生態系裡因為有不同的物種,所以會有「物種多樣性」。一般我們認知的生物多樣性都在「物種」這個層級,比如在這個地方可能有一些鳥類、蛙類、哺乳動物,這些就是它的「物種多樣性」。
「生物多樣性」還有一個層級是「遺傳多樣性」,而且它其實是整個生物多樣性最主要的基礎。「遺傳多樣性」的概念,如圖片中舉例的,同樣的一種蛙類,這是「斯文豪氏赤蛙」,你到野外去,會看到同樣是這種蛙,但有體色有綠色、有很黑色的、有褐色的,你會看到不一樣的地方,這叫「遺傳多樣性」。我們不會把綠色的斯文豪氏赤蛙和褐色的斯文豪氏赤蛙(+認定為)是不同種,為什麼?因為牠們之間可以交配、可以繁殖下一代,牠只是體色不一樣。這時候我們就把它定義為:這是牠們「遺傳」上的差異,而不是不同「種」的差異。這就叫「遺傳多樣性」。
「遺傳多樣性」的概念在森林復育,特別是我們現在講的「天然林復育」就非常重要,重要的原因在哪裡?
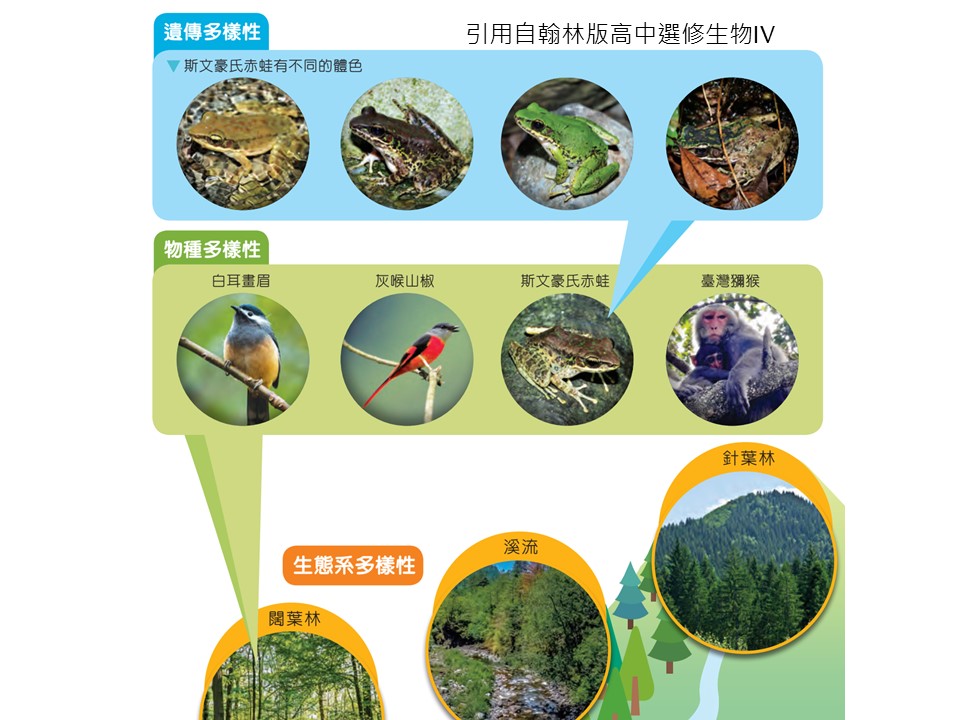
A. 遺傳多樣性是生物多樣性的一環,雖然重要但過去不容易探討
B.遺傳多樣性來自於有性生殖與遺傳變異
C.不同族群出現遺傳變異,稱為遺傳分化
D.遺傳多樣性在生態上的重要性:
i. 確保生物演化的重要基礎。
ii. 增加物種存活的機會。
我們來看第5點遺傳多樣性的「A. 遺傳多樣性是生物多樣性的一環,雖然重要但過去不容易探討」,我們看這張圖。為什麼說「這很重要但過去不容易探討」?因為遺傳分子是很小的,甚至不能用顯微鏡去看,終於後來發展了一些技術。就是過去在新冠病毒covid疫情流行的時候,像是PCR、定序這些名詞大家突然都知道了,因為為了確認新冠病毒是哪一型。這些PCR或定序的技術,在我們科學界可能用了3、40年了,到了新冠病毒爆發社會大眾才知道。這就是我們拿來檢測遺傳多樣性的工具,這是比較近代才發展出來的技術。現在我們可以用這個技術來探討遺傳多樣性,容易探討之後,就會發現遺傳多樣性有很多微妙的地方。
再來,遺傳多樣性第二點,我們看到「B. 遺傳多樣性來自有性生殖跟遺傳變異」。還有「C. 不同族群出現遺傳變異,就稱為『遺傳分化』」。
最後是「D. 遺傳多樣性在生態上的重要性」,遺傳多樣性在生態上有什麼重要,為什麼我們天然林復育要去討論這塊?第i點是確保生物演化的重要基礎,第ii個是增加物種存活的機會。這些我把它綜合成幾張投影片來說明。大家看一下。
「遺傳多樣性」到底有什麼重要?我們來看這張圖。
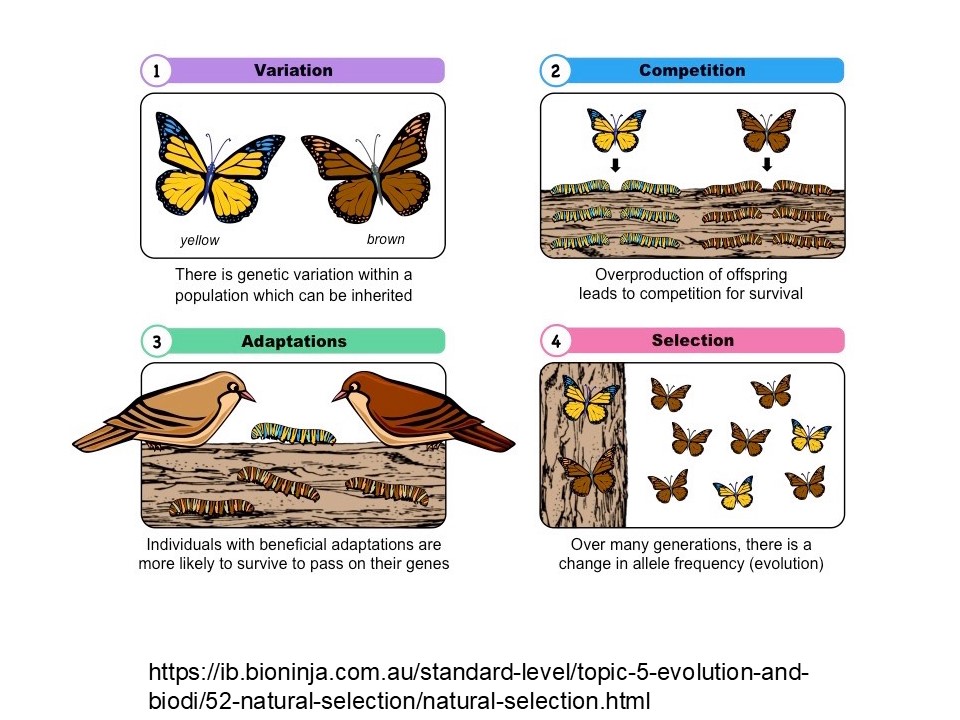
圖1.是表示遺傳多樣性的重要。這是同一種蝶類,牠有兩個顏色,(就像剛才的斯文豪氏赤蛙,牠有不同的體色)。既然是同一種為什麼會出現不同的體色、不同的顏色表徵?因為它的遺傳不一樣,來自於有性生殖、雌雄交配,還有基因突變,因為這樣的結果導致後代體色不一樣。
接下來圖2.,假設牠的幼蟲體色不一樣,一個偏褐色、一個偏綠色,幼蟲都在褐色樹幹上爬。再來圖3.,鳥要過來吃蟲,鳥比較容易看到綠色的蟲,褐色的蟲和樹幹的背景色比較像,所以鳥會先把綠色的蟲吃掉,褐色的蟲有一些雖然被吃掉,但有一些活下來,這叫「適應」。
最後圖4.,你會看到整個蝴蝶族群偏向褐色的居多,因為鳥類篩選了蝴蝶的後代。這蝴蝶牠如果沒有遺傳變異,後代都是同一種的話,可能在一次性的事件裡,牠就滅絕掉了,但我們幾乎看不太到生物會有這種特徵,比如某種生物在某一次重大事件之後造成這個物種徹底滅絕。通常是它的族群被摧毀了一些,或是後代被摧毀了一些,但是有一些活下來。這個例子就是這樣,這是大部分的常態,這就是「遺傳多樣性」的重要性。
所以我們在森林復育要討論遺傳多樣性,因為我們要保證每種樹都具有遺傳多樣性,因為它可以適應未來的環境或環境的變化。你如果不是這樣做,它就很容易在一次的事件裡面全軍覆沒。當然這也不是自然的常態。
這張圖是「臺灣長尾栲的族群遺傳多樣性」。
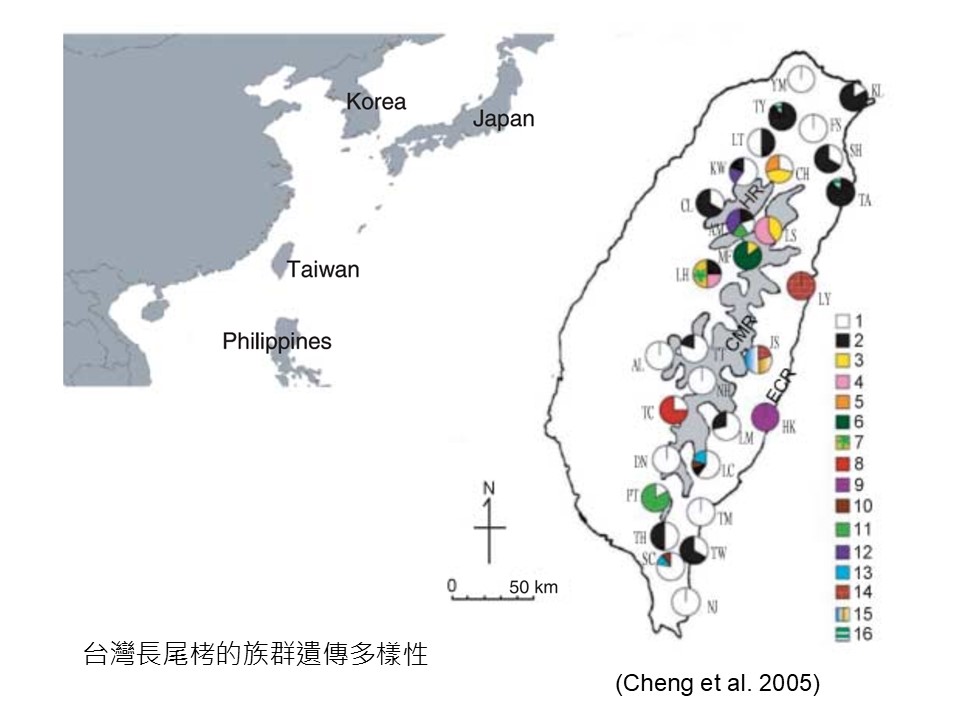
「長尾栲」是廣泛分布在臺灣的一種殼斗科植物。我們的殼斗科植物最常見、(+分布最廣的)就是「青剛櫟」,「長尾栲」是第二分布廣的,而且它在中央山脈從北到最南,以及東部的海岸山脈都有,是一種廣泛分布的殼斗科。
這是鄭育斌我的學長之前做的臺灣長尾栲整個族群遺傳多樣性(研究)。圖上的每一個圓圈代表一個地理族群,顏色差異以及數字是這個研究所定義的基因型。比如白色代表第1型,紅色代表第8型,一些有色彩組合的表示它有2型基因,甚至有3型基因,多彩組合的代表它的遺傳多樣性比較高。
這個研究是要了解長尾栲在臺灣哪裡遺傳多樣性最高,因為他們會認為那裡是長尾栲的「起源中心」,也就是說臺灣的長尾栲族群都會從那邊傳播出來的,這叫「起源中心」。可是我們要探討的不是這個部分,我們是要看,為什麼不同地方的長尾栲會有這些基因型的差異?出現差異的重要性是什麼?
我們看「出現遺傳多樣性的過程」這張圖。
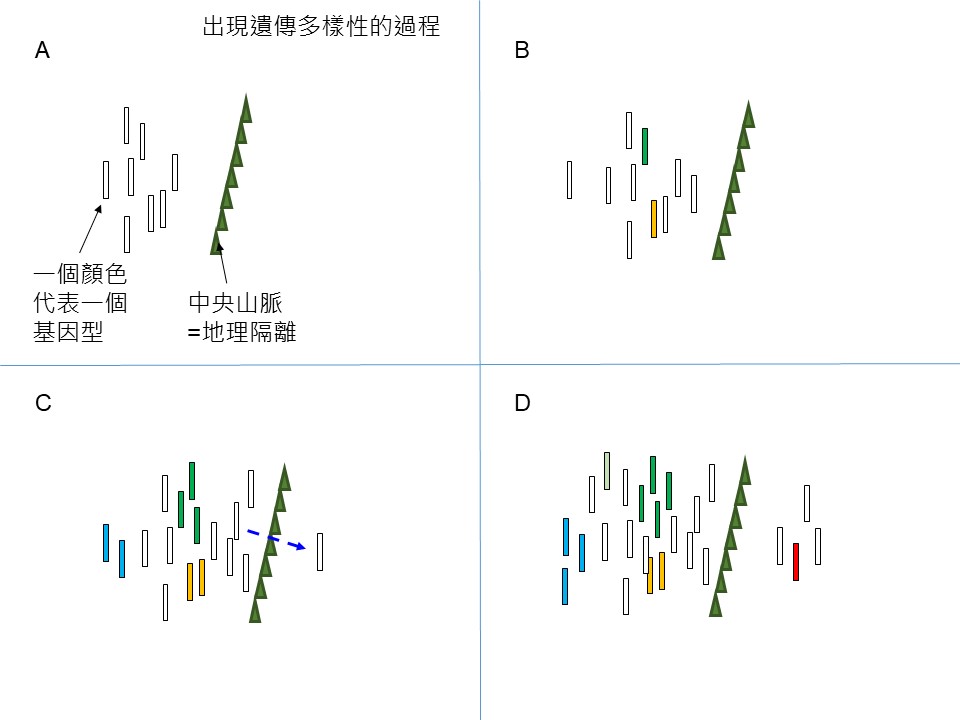
我們假設那個情境,這張圖跟前面長尾栲族群遺傳多樣性的分布圖不是完全吻合的,我只是假設長尾栲的遺傳多樣性的形成是那樣的情境 ,就是:中央山脈我們知道是縱貫臺灣的高山,長尾栲不是高海拔植物,所以它的東、西族群會被中央山脈阻隔,也就是臺灣東部的長尾栲和西部的長尾栲基本上是被中央山脈隔開的。
長尾栲出現遺傳多樣性的過程,可以用這張圖ABCD四個階段來說明。
階段A. 我們假設長尾栲從臺灣海峽的冰期陸橋來到了臺灣西部,它可能是一兩個先鋒先來臺灣定植,開始拓展它的後代,它帶來的基因型非常單純,可能只有一個,假設是圖中白色基因型,這時候因為中央山脈隔開,東部還沒有長尾栲。
下一個階段B,在西部的這些長尾栲因為有性生殖遺傳變異,開始在它的族群裡除了白色基因型,出現了其他基因型(以不同的顏色來表示,如綠色、黃色)。
到了階段C.,可能有某些狀況下,西部的長尾栲可以傳播到東部去,你可以想像這個傳播可能也是先鋒族群過去,比如可能一隻臺灣黑熊吃了以後含在嘴巴,越過中央山脈到那邊吐掉,把它的果實帶過去(笑),假設這樣的情境,牠可能帶了某個基因型到東部來。這時候西部的基因型就越演化越多,因為後代不斷地生殖,遺傳變異(綠、黃、藍)。
再接下來,階段 D.,我們會看到東部的長尾栲的後代也產生了遺傳變異(紅),西部當然繼續遺傳變異,東部也開始出現。我們可以預期:東部後代的遺傳變異和西部的遺傳變異會不一樣的。遺傳變異一樣的機率太小了,因為遺傳變異是隨機出現,既然是隨機出現,它要完全一樣的機率就很小。就好像人類 ,在臺灣2,300萬人裡說不定哪天會找到一個跟你長得很像的人,但跟你沒有血緣關係,這是很小的機率,你不會遇到1,000個人、100個人,都長得像你,就是這個意思。
所以東部、西部這兩個族群的後代各自出現遺傳變異,但是它們之間是沒有基因交流的,所以是各自遺傳各自演化,所以它們出現的遺傳變異是不同的,正常狀況是這樣。這就是解釋「臺灣長尾栲的族群遺傳多樣性」那張圖。
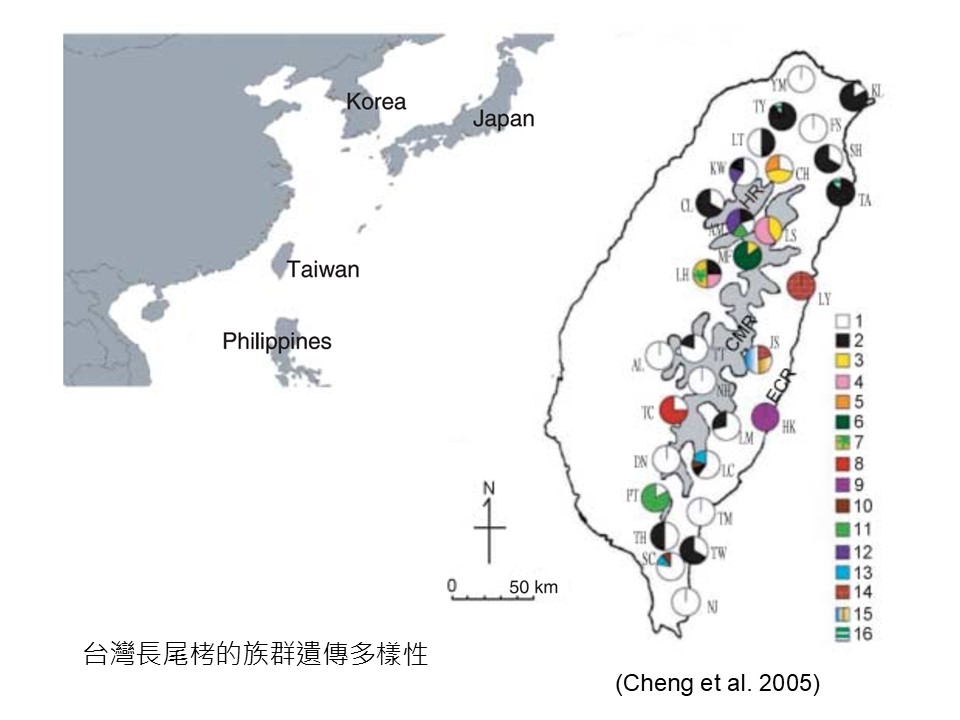
你從這張圖可以看到長尾栲最主要的基因型是白1,因為它幾乎廣布在所有族群,除了少數族群沒有白1之外,其他都有,所以白1它一定是最基本的基因型。然後它開始到處傳播之後,後代開始出現遺傳變異,圖上的不同色塊的,就是代表白1的後代出現了遺傳變異。
經過很久之後,這可能是幾十萬年的尺度,它慢慢廣布到臺灣全島,出現很多不同基因型,最早來臺灣的可能就是白色這種基因型。
在這張圖也可以看到,東部的基因型很不一樣,海岸山脈在花蓮這邊有一個獨特的基因型紅14,這是西部沒有的;還有一個藍13的基因型只在東部而已,西部也沒有。所以(+遺傳變異的產生)大概是這樣的過程。
當這個物種開始出現族群,它開始傳播、演化、它就會出現「遺傳變異」,這就是「族群的『遺傳分化』」。
這也就關聯到最後一個主題6「在地苗木」,關聯到「遺傳多樣性的重要性」。為什麼我們在森林復育不要使用非在地的苗木,「要盡量用在地苗木」?原因在這裡。
6.在地苗木:
A. 復育時只考慮物種,雖符合物種層級的多樣性考量,但不符合遺傳多樣性的原則。
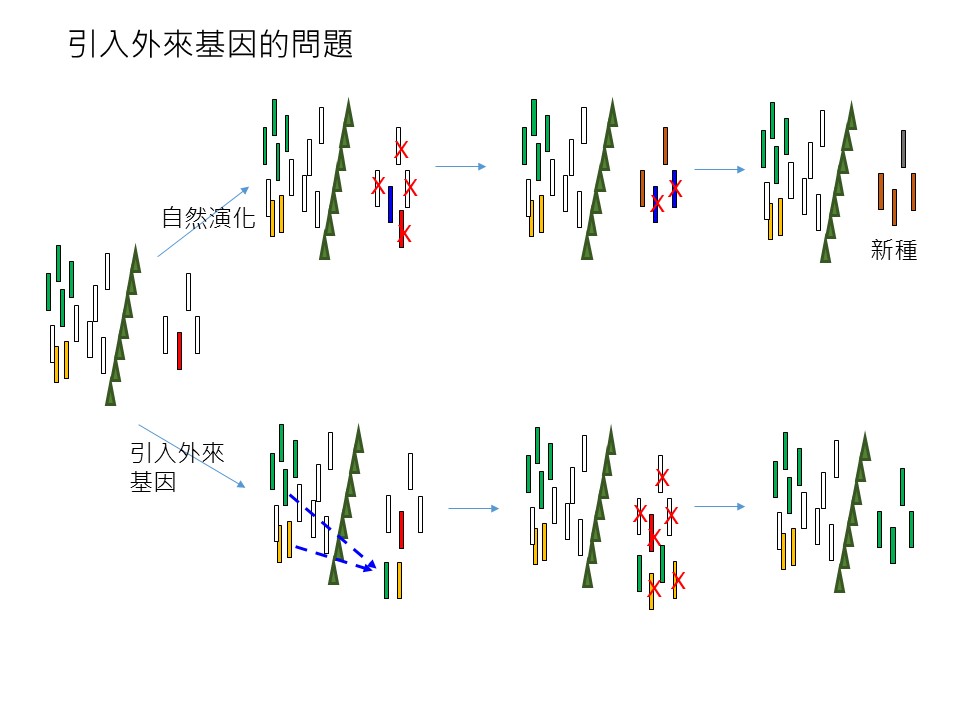
我們延續前一節的「長尾栲」的狀況來討論,在「引入外來基因的問題」這張圖的最左邊是「長尾栲已經產生遺傳變異」的狀況。
假設「長尾栲」自然演化,它應該是要走圖的上方「自然演化」這條演化途徑,這條途徑我解釋一下:隨著時間過去,它不斷地有性生殖,繁殖產生下一代。你會看到這些遺傳基因型一直保留下來,山脈的西側跟東側的基因型都有保留下來,在東側有「打叉」的,是代表經過某個特別的事件,這事件也許是東部有一個很不一樣的氣候出現,假設是長期的焚風,在西部活得好好的這些基因型白色的族群,它到了東部它不適應焚風這種太高溫的環境,它就死掉。「打叉」代表它死掉被淘汰了;但是可能某些在繁殖的基因型,比如藍色的,它是適應焚風的環境,它活了下來。假設沒有遺傳多樣性的話,(+這些長尾栲)就是全軍覆沒了,但有遺傳多樣性的,可能有些後代可以在這個篩選過程中存活下來,比如這藍色基因型的。這就是「遺傳多樣性」的重要性。
接下來,可能持續地不好的環境不斷地出現,你也可以預期藍色基因型的,它也會再產生一些遺傳變異的後代,這些後代可能又可以活下來。所以最終因為中央山脈隔離的結果,這些後代變異已經跟原先的祖先(白色基因型)很不一樣了,就演變成所謂的「新種」。
我們假設,這是一個假設,不是事實:東部有一種「灰背櫟」的植物長得很像長尾栲,長尾栲沒有毛,灰背櫟葉背都是毛,樣子很像長尾栲。我再強調一次,這是假設,我假設「灰背櫟」搞不好是這個狀況出現的,我是舉例。所以這種叫做「新種」,意思就是「遺傳變異」到最後和「母族群」、「母種」本身差異很大的時候,我們會把它定義為一個「新種」。它就要靠這樣的過程才會出現新種。
所以我們的「物種多樣性」是怎麼來的?是「遺傳多樣性」來的。生物經由演化不斷出現遺傳多樣性,然後自然的篩選、淘汰,「遺傳多樣性」一直循環,最後「物種多樣性」就出來了,也就是從「一個種」變成「很多種」。
所以我們如果能夠確保一個族群的遺傳多樣性,就可以確保一個物種的演化,可以確保物種多樣性。所以自然演化就是圖上方的這條途徑。
圖下方是「引入外來基因」這條途徑。什麼是「外來基因」?先前我們提到,如果我們進行臺中大肚山復育要種「三斗石櫟」,大肚山低海拔,但因為宜蘭平地也有「三斗石櫟」,我們去宜蘭平地找一些種源(+過來種),這樣行不行?當然不好。宜蘭和臺中位置相距很遠,一個在東部一個在西部。從長尾栲的例子你可以知道,應該宜蘭的三斗石櫟和大肚山應該有的三斗石櫟,基因型會差很多。
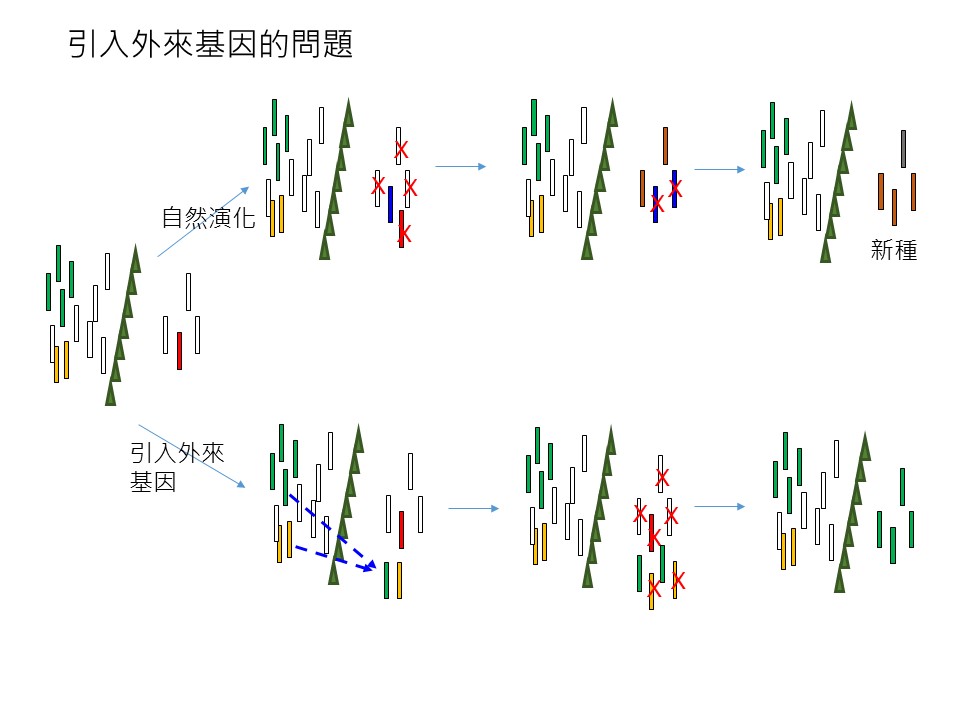
如果你把宜蘭的「三斗石櫟」引過來臺中種會怎麼樣?可能就是圖下方「引入外來基因」這條途徑:本來都在西部的(基因型綠、白、黃),東部也有一些(綠、白、黃),我們現在在東部要來復育這個種類,比如在花蓮溪口,本來預期應該要有一片長尾栲為主的森林,但現在沒有。我們在造林的規劃裡就說,要在那邊種一些長尾栲,但是我們從西部去拿種源,這時候我們會把某些西部的基因型帶過來。但是本來在東部山區,比如花蓮溪附近的海岸山脈或山區本來就有一些長尾栲,這時候你又從西部帶過來東部沒有的基因,東西部這兩個一定會雜交,雜交之後會把東部好不容易演化出來的特別的基因消除掉,會被西部的基因「均化」掉了,抵銷掉了,所以它會變得比較像西部的族群,東部這些特別的基因可能就不見了,而西部過來的基因還保留在東部這邊。
這樣的後果就是,第一個就是「遺傳多樣性降低」了,接下來還有可能會發生,假設「環境的篩選」它可能把某些基因型除掉了,西部你移過來的剛好是適應東部這個環境,結果變成東部都是你從西部帶過來的基因型,變得很單純,然後東部的基因型它就完全沒有了,它就被人為干擾了,遺傳多樣性的變化就被你擾亂掉了,最重要的是-它形成新種的能力-就被你往回推。本來東部的族群它是隨著時間過去,可能它越來越有機會變成新種的,你現在把它往回推,也就是倒退,變得很不容易出現新種,那麼物種多樣性就會降低。
提問:做這些事的人他自己看不到他干擾了演化的後果?
答:當然看不到,而且最重要的是他沒有概念。因為主要做這些(復育?種樹?)的人我們都沒有受過這個教育。我剛才提到,我們現在對遺傳多樣性的檢測可能是三、四十年來才有的技術,技術剛出來的時候臺灣只有非常非常少數的人在做,現在是很多人在做了,可是在當時沒有,所以大部分人不曉得。他就會覺得,反正我要種這個種類,我只要種就好了,為什麼我還要在意我從哪裡拿到的種源呢?
但是大家看看協會的作法,我們在大肚山做復育,我們(+採種的來源)是大肚山優先,接下來是鄰近山區(比如火炎山、大坑、八卦山),從來不會去考慮到東部的物種,原因就是在「維持遺傳多樣性」。我們在做這件事的時候才能確保這個物種演化的未來。你沒有做得更好,也不要把它推向一個更壞的情況,只要你引入外來基因就是在走回頭路了,就是在摧毀物種的遺傳多樣性,在摧毀物種的未來。
提問:之前上課的時候,蘇老師您有講過烏心石的例子。
答:是類似那樣的例子。烏心石的造林就是一種很麻煩的,同一種的烏心石到處被亂種,甚至拿蘭嶼的烏心石來臺灣種,連不同種也加進來。
我剛才講的,我們在森林復育,在自然林這一塊,沒有人這樣做,這是比較可惜,因為做這個需要滿大的經費,要滿有概念的人進來做。
蔡:剛才講到經費的部分,林管處一棵樹苗的採購經費,一棵是30塊,1萬棵就是30萬。就我們協會這幾年在運作的狀況下,我去算成本,一棵苗的整個成本是要1,000塊,但是這已經高到林管處的主計都覺得不可思議,比市價的苗木都還要來得貴。為什麼我們算得出來?因為我們投入運作的成本,一年可以出多少苗,整個運作的狀況下來,所有的人事成本加起來,一年就是出1萬棵。所以1棵苗就是要1,000塊,但是社會還沒辦法接受這個成本這麼高。
蘇:這是在行政面上的。因為他們過去是用比較造林的方式,造林有所謂的「採種母樹」,採種母樹都是那種產量非常高的、他們認為很健康的(+個體)。所以像「檜木」他們的採種母樹非常單一,「長尾栲」的採種母樹非常單一。比如這個林管處是負責長尾栲的,他可能就從那一棵母樹採了一大堆,然後這些後代分送到有需求的林管處,就調苗種到那邊。林管處造林的方式就是一直在走「引入外來基因」這條路。為什麼林管處他們的苗可以那麼便宜,因為他們都從單一的採種母樹來,沒有在意遺傳多樣性,這個就很簡單,可以用大批的、大規模的方式去生產。
B. 使用外地苗木,等於是引入外來的遺傳基因,會破壞自然演化的過程。附帶也可能會引入外來疾病(如白輪盾介殼蟲)。
我有其他的例子來說明遺傳多樣性被摧毀的狀況。

這個是基改作物的抗殺草劑基因,為什麼基改作物要放抗殺草劑的基因?因為要避免作物被殺草劑除掉。在國外像美國那種很大的土地,都是用機械式耕作。它用殺草劑下去除草的時候,例如年年春這一類的,是所有的植物都摧毀掉,因為這類殺草劑會阻斷植物的生化反應,每一種植物全死。所以他(?)為了要讓殺草劑有作用,他會在自然界去測試。比如在自然界物種很多,去大規模地使用殺草劑,用了之後發現有一兩棵植物竟然活在那裡好好的,其他死光光了,那就是遺傳變異,就是(還活著的)這個植物體內有一種基因可以產生抗殺草劑的蛋白質。所以這個植物沒死,其他植物都死了。
(?誰)他就去找這個(+抗殺草劑)基因轉殖到作物,比如大豆或是小麥,轉殖進去之後你所種的作物全部都具有抗殺草劑基因。所以他再來施用殺草劑,只會把雜草殺死,作物不會死,因為這作物具有抗殺草劑的基因。所以現在的基改主要是在做(+抗殺草劑作物)這個部分。
久而久之,他們發現,用(抗殺草劑作物)這一招種過的田,發現了某些雜草除不掉。比如圖(a)這個田是加拿大蓬、(b)是一些我們常見雜草、(c)是某一種莧 。(+使用)這個殺草劑過去以後,不只作物殺不死,連雜草也殺不死。他們就去找原因,發現這些殺不死的雜草從這些(+基改)作物意外地獲得了抗殺草劑基因。但(+雜草和作物)它們是不同種的植物,這基因怎麼來的?這個稱為「基因滲入」,可能有一些病毒去感染這個(+基改)作物之後,從這個作物身上獲得抗殺草劑基因在這病毒體內,這個病毒繁殖出來之後又去感染旁邊的這些雜草, 所以這些雜草就從病毒裡面獲得(+抗殺草劑)基因。
其實我們的人工基因轉殖也是模仿自然界病毒的這種操作,結果這病毒就是在大自然做基因轉殖,把它轉到雜草,所以我們把這種雜草稱為「超級雜草(Super weed)」,抗殺草劑基因是殺不死它的。
這就是我們講的,你在違反生物多樣性。
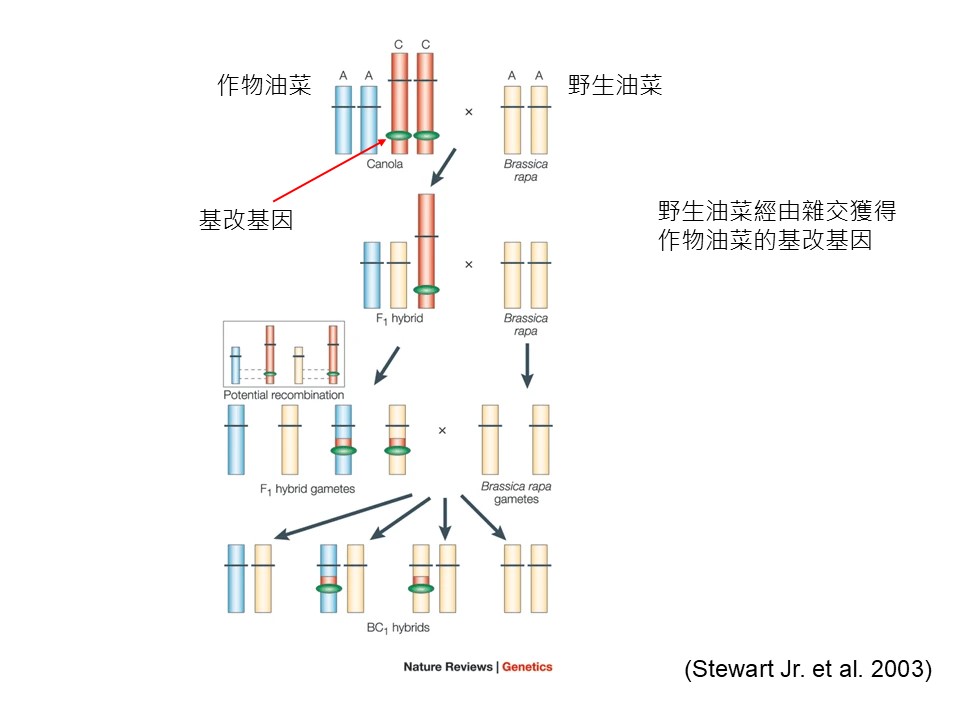
另外一個例子是「作物油菜」,它也是被殖入基改基因。
「作物油菜」和「野生油菜」有不同的染色體的狀態,他們把它殖入在不同染色體上。一般來講,「作物油菜」和「野生油菜」是不會雜交的,因為染色體不一樣。染色體不一樣在有性生殖配對時,染色體會異常,後代沒辦法成形。因為作物油菜跟野生油菜染色體的配置是不同的,所以他們(+研發人員)不擔心基因會傳到野生油菜去,結果沒想到竟然還是傳過去了。
結果他們去研究(+基因移轉的?)路徑發現,這個染色體在作物油菜裡面進行染色體的互換,(染色體互換大家再另行參考高中生物)。(+作物油菜)只是純粹在它的細胞裡面做染色體的互換,就把抗殺草劑基因轉到可以跟野生油菜配對的染色體上面,於是它們就可以正常去配對,所以野生油菜的後代就獲得抗殺草劑基因。
所以電影侏儸紀公園有一句話說:「物種會自己找路。生物會自己找它的路。」這(個例子)就是自己找它的路。我們一直認為我們可以用人類的方式可以去阻斷它的生殖,實際上是做不到的。這(+個例子)是實際上發生的事情。
如果我們去引入外來基因,我們就會用人類的方式把物種演化的自然能力摧毀掉。表面上這物種是還在這邊的,但你只是看到現況。未來的話,這物種未來演化的能力被你摧毀掉了。所以我們最好是用在地的苗木。
C. 討論:復育上是否一定不能引入外來基因?
從前一節的討論,我們知道引入外來基因是不好的,但是我們在做天然林復育的時候,一定不能能引入外來基因嗎?或者有些狀況是可以做的?

蔡:如果(+這個物種)是區域滅絕的話,就不得不這麼做了。
蘇:那要定義一下「區域滅絕」。
蔡:比如(+這個物種)過去標本的紀錄在大肚山有,但是現在怎麼找都沒有了,在周圍的火炎山、八卦山、大坑也都沒有了,我們可能就要在更往北到苗栗或雲林那邊找進來。
蘇老師補充說明:「物種滅絕」有幾種層次,一種層次叫做「滅絕」。「滅絕」是最慘的,就是地球上再也沒有這個物種了。比如澳洲的「袋狼」,牠再也沒有出現在地球上;又例如恐龍是滅絕的,這 一類的生物。
有一種層次是「野外滅絕」,意思是在野外看不到了,都沒有野生族群,但是在人類圈養的環境裡面它還存在。例如「銀杏」就是野外滅絕的物種。
「區域滅絕」是滅絕的層次裡面程度比較輕微的,當然還是不好。「區域滅絕」的意思是在我們設定的某個地理範圍內,它本來有的,但現在都沒有了。例如我們很多的水生植物常常出現區域滅絕的狀況,但是在其他地方還有這個物種。
「區域滅絕」還提供一個可能性是,我們還是有其他種源可以拿回來復育,可是這時候你引入的一定是外來基因,但是因為這個物種在這邊已經完全都沒有個體,你不得不做這件事,所以有時候引入外來基因有它的必要性。
蘇:剛剛是說區域滅絕的情形,我再進一步問,假設這個地方還有這個物種,我們能不能引入外來基因?
蔡:當這個物種它繁衍的能力已經很弱了,因為它的基因弱化了。這個是在大肚山很明顯的例子,就是這物種它的小苗的狀況都比其他區域的小苗的健康狀況差異很大。會不會是這樣?
蘇:就是當這個物種它在這個地方它表現的生長已經不是很良好的時候,我們可能有需要適度地引入外來基因。我們有一種概念是:遺傳多樣性高的地方,它這個族群會比較健康。如果遺傳多樣性很低,甚至基因型是單一的,代表都是「近親交配」。「近親交配」在生物裡面常會講一個現象叫「近交衰敗」。近親交配後,它的後代會變得很弱,比如人類很明顯,如果是近親交配,血緣關係很近的人結婚有後代的話,後代常有遺傳疾病出現,這就是一種「近交衰敗」的現象。
所以確實我們在復育上,如果這個地方我們要復育的物種它已經面臨遺傳多樣性很低的狀況時,有時適度地引入鄰近地區的同種生物、不同基因型,可以適度地讓這個族群變得比較健康。
因為你要想想看,在大自然裡,這個物種它本來遺傳多樣性是很高的,它之所以遺傳多樣性會很低表示是過去我們的破壞太嚴重(所造成),這時候你從別的地方引入一些不同的基因型過來,有它的必要性。不然你可能努力了半天還是沒有用。
這在國外的野生動物復育還滿常用的。比如有某種瀕臨滅絕的動物,它在這地方的族群已經遺傳多樣性很低了,我們雖然已經救活牠,設立保護區了,牠的數量也達到某個程度,可是遺傳多樣性還是很低。如果我們放任不管讓牠自己繁殖,(近交衰敗),最後還是會滅絕。所以這時候從比較遙遠的地方去引入一些個體來跟牠交配,讓遺傳多樣性拉高,這個(+方法)已經有在做。所以在某些情況我們可能有須要去實施,但是通則是最好先不要做,除非確定。
提問:臺灣的「雲豹」已經滅絕,可不可以從其他的國家再引進跟臺灣不同亞種的雲豹進來。這件事情在脈絡上面跟您的例子是一樣的?
答:對,脈絡上是一樣的,但是雲豹的問題比較麻煩。植物的好處就是植物不會吃別的(生物)了。
D.在地苗木的確認,由採種母樹開始。確認採種母樹的原生性,請參考本課程第2主題有關”剔除外來種與非當地的臺灣原生種”的部分。
最後我們來談確認採種母樹這件事,因為我們要培育在地種苗。
我們之前的課程已經有跟大家討論過,我們再回顧一下。「在地苗木的確認,由採種母樹開始。」最好的方式還是採在地的母樹去培育,得到在地的苗木來做復育。

我們要怎麼去確認,在調查或採種的地方看到的這些我們認為的母樹它是原生的?有幾個原則可以來判定。第一個原則:「剔除外來種與非當地的臺灣原生種」。只要是「外來種」就剔除,臺灣的外來種當然不可能是當地原生的,其次有一些「非當地的臺灣原生種」,要判斷「非當地的原生種」就比較麻煩,但是可能有一些蛛絲馬跡(可以判斷)。
如果是「非當地的臺灣原生種」通常會是「造林」或「景觀」需求而種的。造林種下的,會是大量的、同齡的形式出現。例如我們常看到野外的相思樹很多都是這樣,你去看都沒有小苗,都是大樹,樹齡也差不多,這應該就是造林的。
為了「景觀」種植的物種,那種情況通常會零星地出現在步道邊,如果它自己能自然下種的話,在森林內部會缺乏(它的)族群,因為從外地引過來的這些景觀樹種它通常不適應當地的環境。當地的森林因為長久演化,在能活在裡面的物種都是適應那個環境的,一般我們引入外來的植物通常它沒辦法長在那種環境。所以是景觀樹自己天然下種的情況,通常會出現在步道邊、路邊而已,它不會進到森林裡面。如果(物種)是這種分布型的,你也可以先把它排除掉。
我們在進行森林復育應該是先用比較保守的方式去做,我們確定沒問題的先做。如果真的材料(物種數)很少,才去往外擴張找我們不確定的。如果我們確定的(物種)已經相當足夠的話,其實問題就很小了,因為我們光要處理這些已經確定的物種,就要花很大的心力了。
以上是「種子特性與遺傳多樣性」的課程內容。
-本章結束-